题目列表(包括答案和解析)
(十)尿的生成过程
尿是在肾单位和集会管中生成的。其生成的基本过程为:①肾小球的滤过作用;②肾小管与集合管的选择性重吸收作用;③肾小管与集合管的分泌和排泄作用。
血浆通过肾小球的滤过作用生成原尿;原尿通过肾小管和集合管重吸收和排泌作用生成终尿。
1.肾小球的稳过作用
血液流经肾小球时,除血液有形成分及血浆中大分子蛋白质外,其余的可透过滤过膜,进人肾小囊内形成原尿。
肾小球滤过率指单位时间内两肾肾小球滤过的滤液量(即原尿量),大约是125ml/min,两侧肾脏24小时的滤过量约为180L。
决定及影响肾小球滤过作用的因素包括:
(1)滤过膜是指血浆通过毛细血管进入肾小囊所跨越的膜性结构,是肾小球滤过作用的结构基础。膜上有大小不等的孔道,血细胞和大分子物质(如蛋白质)不能通过孔道法过。滤过膜还有负电荷,能阻止带负电荷的物质通过滤过股,可以把滤过膜看作选择性滤器。病理情况下,滤过膜受损,通透性增加,尿中可出现蛋白质,即蛋白尿,甚至出现红细胞,即血尿。正常成人滤过膜的总面积约为1.5m2,面积减少可使滤过率降低,出现少尿或无尿。
(2)有效滤过压是使血浆通过滤过膜而滤出的净压力,为肾小球滤过作用的动力。
有效滤过压=肾小球毛细血管压一(血浆胶体渗透压+肾小囊内压)
肾小球毛细血管血压是推动血浆滤出滤过膜的力量,而血浆胶体渗透压和囊内压是对抗滤出的力量。这三种力量的变化则是影响有效滤过压的因素,即可影响原尿生成量。肾小球毛细血管血压主要取决于全身动脉血压的高低和出入球动脉的口径。当动脉血压在80-180mmHg时,通过肾血流量自身调节,肾小球毛细血管压变化不大,有效滤过压变化也不大。当动脉血压低于80mmHg时,肾小球毛细血管血压降低,使有效滤过压降低,引起少尿或无尿。血浆胶体渗透压和囊内压在生理情况下变动不大,因而对有效滤过压和原尿量的影响很小。但在静脉快速注入较大量的生理盐水时,可使血浆胶体渗透压降低而增加有效滤过压。发生尿路结石、肿瘤压迫引起输尿管堵塞时,可使囊内压升高而降低有效滤过压。
2.肾小管和集合管的重吸收作用
每昼夜生成原尿量约180L,而终尿量却只有1L-2L,为原尿量的1%,这是由于原尿经过肾小管和集合管时,将其99%的液体又重吸收回血液。
原尿流经肾小管时,其中某些成分重新返回血液的过程称为重吸收。
近曲小管是重吸收的主要部位。重吸收有两种方式,即主动重吸收和被动重吸收。K+、Na+在肾小管吸收是主动的。Na+重吸收所造成的管内外电位差,引起Cl-的被动重吸收。
只有近曲小管有重吸收葡萄糖的能力,但其重吸收能力有一定限度,超过其限度就不能全部重吸收,在尿中出现糖称为糖尿。
当尿液中开始出现糖时,血浆中糖的最低浓度称为肾糖阈,正常人肾糖阈为160-180mg%。
水的重吸收是被动的,在近曲小管和髓襻主要是伴随溶质的吸收而吸收。在远曲小管和集合管的重吸收还要受激素影响,其重吸收率决定体内的含水量,当体内水分多时,水重吸收减少,体内水分少时,则重吸收增加,以保证机体的水平衡。水的重吸收率在99%以上,重吸收率改变1%,就会引起终尿量成倍的变化。
3.肾小管和集合管的分泌和排泄功能
肾小管上皮细胞将血液中某些物质排入小管液中的过程称为排泄;肾小管上皮细胞将自身代谢产生的物质排入小管液中的过程称分泌。分泌和排泄都是通过肾小管细胞进行的,分泌物和排泄物都进入小管液中,所以通常对两者不作严格区分。
(1)K+的分泌:主要由远曲小管、集合管分泌。由于Na+的主动重吸收,使管内为负,管外为正的电位差,K+就顺着这一电位差被动扩散入小管液,形成K+-Na+交换。
(2)H+的分泌:肾小管各段和集合管上皮细胞都能分泌H+,这是依赖H+泵的主动运转过程。分泌到管腔液中的H+,可与管腔液中的HCO3-结合成H2CO3,H2CO3分解成 H2O和CO2,此CO2通过细胞膜进入肾小管上皮细胞内,在其中的碳酸酐酶的作用下,CO2与H2O结合成H2CO3,它立即离解成H+和HCO3-。H+分泌到管腔液中,由于电位差而促进Na+的吸收,形成Na+-H+交换。细胞内形成的HCO3-,由于膜的特性只能通过小管上皮细胞的基底膜一侧扩散入细胞间液最后吸收到肾小管毛细血管中,而进入血液循环。因此H+的分泌既可促进Na+的吸收,又可促进HCO3-的吸收,而 NaHCO3是人体的碱贮。这种排酸保碱作用对调节体内的酸碱平衡具有重要意义。
K+-Na+交换与H+-Na+交换之间有相互竞争作用。当K+-Na+交换增多时,H+-Na+交换减少;反之,当机体酸中毒时,H+生成增多,H+-Na+交换加强,从而抑制了K+-Na+交换,所以,酸中毒时,常有血K+增高的现象。
(3)NH3的分泌:氨是脂溶性物质,可扩散到小管液中与H+结合成NH4+,降低小管液的H+浓度,有利于H+的再分泌。
(九)消化液的成分及作用
1.唾液及其作用
唾液是唾液腺分泌的无色近中性(pH为6.6-7.1)的液体。正常成人每日分泌量l-1.5L。唾液含有99%的水,还有唾液淀粉酶、粘蛋白、溶菌酶及少量的Na+、K+、Cl-等。
唾液的作用:①湿润和溶解食物,使之容易吞咽及引起味觉。②唾液淀粉酶可将淀粉分解为麦芽糖。此酶受氯离子激活,其作用用最适的pH近于中性。由于食物在口腔停留时间很短,淀粉酶的作用不能充分发挥,故细嚼慢咽,能增加淀粉的分解。③清洁保护口腔。唾液经常分泌,能湿润口腔粘膜,清除口腔内的残余食物或异物。溶菌酶有一定的杀菌作用。因此,对唾液分泌过少的患者,应当注意保护口腔。
2.胃液及其作用
胃液是胃腺和胃粘膜上皮细胞分泌的无色、强酸性(pH为0.9-1.5)的液体。正常成人每日分泌量约1.5-2.5L。胃液有大量水分、盐酸、胃蛋白酶、粘液、内因子和Na+、K+、Cl-等。
(1)盐酸:盐酸由胃底腺的壁细胞分泌。通常所称的胃酸即指盐酸。胃液中的盐酸,大部分为游离酸;小部分盐酸与蛋白质结合为盐酸蛋白盐,称结合酸。两者酸度的总合称为总酸。
盐酸的作用:①激活胃蛋白酶原,提供适宜胃蛋白酶活动的酸性环境;②使食物中的蛋白质变性,容易被水解;③有杀菌作用;④进入小肠后,可促进胰液、胆汁、小肠液的分泌,并有利于小肠对铁、钙的吸收。胃酸不足时,胃肠的消化能力随之减弱,细菌容易生长繁殖。胃酸过多,会侵蚀粘膜,可能是胃、十二指肠溃疡的病因之一。
胃粘膜有防止H+从胃腔侵入粘膜内,又能防止Na+从粘膜内透出的作用,称为胃粘膜屏障。胃粘膜屏障的存在,使胃粘膜与胃腔之间维持着悬殊的H+浓度差。这样既能保持胃腔内盐酸有一定浓度,又能防止盐酸侵蚀胃粘膜,具有胃的自身保护作用。胃粘膜受到细菌侵袭、缺血、缺氧,以及与较高浓度的酒精、醋酸和阿司匹林等物质接触后,可使粘膜屏障受损,大量H+迅速向粘膜内扩散,破坏胃粘膜细胞,导致溃疡病。
(2)胃蛋白酶:由胃底腺主细胞分泌出来的是无活性的胃蛋白酶原,它在盐酸的激活下成为胃蛋白酶。已被激活的胃蛋白酶也可激活胃蛋白酶原。胃蛋白酶在强酸环境中能使蛋白质水解为眎、胨和少量多肽、氨基酸。
(3)粘液:是由胃腺粘液细胞和胃粘膜上皮细胞共同分泌的,其主要成分是粘液蛋白。粘液蛋白具有较高的粘滞性和形成凝胶的特性,分泌后覆盖在冒粘膜表面,形成一凝胶保护层,具有润滑食物和防止粗糙食物对鲁粘膜的机械性损伤。还可降低H+在粘液层中的扩散速度。
(4)内因子:内因子是由胃底腺的壁细胞分泌,它能够与维生素B12结合成复合物,使之不受破坏,并促进其吸收。因此,缺乏内因子时,会产生巨幼红细胞性贫血。
3.胰液及其作用
胰液是胰腺分泌的无色、碱性(pH为7.8-8.4)的液体,正常成人每日分泌1-2L。胰液中含水、碳酸氢盐和多种消化酶及Cl-、Na+、K+等。
(1)碳酸氢盐:有中和进入十二指肠的胃酸,保护肠粘膜免受胃酸侵蚀的作用。还能形成适宜于小肠内多种消化酶活动的pH环境。
(2)胰淀粉酶对生熟淀粉均有催化作用,其催化效率高,可将淀粉分解成麦芽糖和葡萄糖。
(3)胰脂肪的:催化脂肪分解成甘油和脂肪酸。
(4)胰蛋白酶和糜蛋白酶:这两种酶刚分泌出来为无活性的酶原。胰蛋白酶原受酸、肠激酶及胰蛋白酶激活;糜蛋白酶原受蛋白酶的激活。胰蛋白酶和糜蛋白酶均可以催化蛋白质分解成眎、胨。两种酶共同作用,可以将蛋白质分解成多肽和氨基酸。
胰液含的消化酶种类多且较全面,是消化力最强的消化液。若胰液分泌过少或缺乏,将出现消化不良,食物中的脂肪和蛋白质不能被完全消化和吸收。
4.胆汁及其作用
胆汁是肝细胞分泌的,不含消化酶的消化液。
(1)胆汁的分泌、贮存和排放:肝细胞不断分泌胆汁,成人每日约分泌800-1000ml。在非消化期,胆汁流人胆囊贮存。在消化期,随着胆汁的排放,肝细胞分泌的胆汁可以沿肝胆管道直接进入小肠。进食之后的消化期,经过神经及体液调节,肝细胞分泌汁增多,胆囊的胆汁排放。胆汁排放时,胆囊收缩,壶腹括约肌舒张,肝胆汁与胆囊胆汁均排入十二指肠。
(2)胆汁的成分和作用:胆汁是浓稠有苦味、有颜色的液体,颜色决定于胆色素的种类和浓度。肝胆汁为金黄或橘黄色,呈弱碱性(pH约为7.4)。胆囊胆汁因部分水分和碳酸氢盐被胆囊吸收而被浓缩,颜色较深、pH约为6.8。胆汁主要含胆色素、胆盐、胆固醇、卵磷脂及多种无机盐。
胆汁中与消化、吸收有关的成分主要是胆盐。胆盐是各种结合胆酸形成的钠盐的总称。胆盐的作用:①一定量的胆盐能提高胰脂肪酶的活性,加速它对脂肪的分解作用。②使脂肪乳化成极小的微粒,增加脂肪与脂肪酶的接触面,利于消化。胆汁中的卵磷脂、胆固醇也有相同的作用。③胆盐可以与脂肪酸结合形成水溶性复合物,促进脂肪酸的吸收,同时也促进脂溶性维生素的吸收。
肝脏、胆道患病者,胆汁分泌减少或排放受阻,会出现脂肪的消化和吸收不良,以及脂溶性维生素吸收障碍。
5.小肠液及其作用
小肠液是弱碱性(pH约7.6)液体,成人每日分泌约1-3L。小肠液中除肠激酶外,还有淀粉酶、肠肽酶、二糖酶(包括麦芽糖酶、蔗糖酶和乳糖酶)分别将淀粉水解为麦芽糖,多肽水解为氨基酸,二糖水解为单糖。有人认为除肠激酶是小肠分泌的之外,其他消化酶是由小肠上皮细胞脱落破裂进入小肠液。此外,大量的小肠液能使消化产物稀释,有利于小肠粘膜的吸收。
6.大肠液及细菌的作用
大肠粘膜的柱状上皮细胞和林状细胞分泌大肠液。大肠液富含粘液和碳酸氢盐,是碱性(pH为8.3-8.4),具有润滑粪便、保护粘膜的作用。
大肠内的细菌还能利用肠道内某些简单物质合成B族维生素与维生素K,可被吸收供人体。因此,长期使用肠道抗菌药时,要注意补充上述维生素。
7.糖、脂肪和蛋白质的吸收形式和途径
(1)糖类的吸收
一切糖类都必须被消化成单糖才能在小肠被吸收。小肠内的单糖主要是葡萄糖。单糖的吸收主要通过毛细血管进入血液,经门静脉到达肝脏,而后在肝脏内贮存和分布全身。
(2)脂肪的吸收
脂肪消化产物脂肪酸、甘油一酯和胆固醇,与胆盐形成脂肪微粒到达小肠微绒毛上后,脂肪酸、甘油一酯、胆固醇进入粘膜细胞,胆盐留在肠腔。
长链脂肪酸(15个碳原子以上)和甘油酯在肠上皮细胞内重新合成甘油三酯,并与载脂蛋白合成乳磨微粒,经淋巴吸收;中、短链脂肪酸(15个碳原子以下)直接经毛细血管吸收。
(3)蛋白质的吸收
食物中绝大部分蛋白质在小肠内被分解为氨基酸而被吸收入血液。
水、无机盐和维生素不经消化而被小肠直接吸收入血液。胃也能吸收水分。
(八)气体在血液中的运输
O2和CO2在血液中运输的形式有两种,即物理溶解和化学结合。物理溶解的量虽很少,但很重要。因为气体必须先通过物理溶解,而后才能化学结合;在化学结合或解离时,又须通过物理溶解而扩散。
1.氧的运输
(1)物理溶解:物理溶解的量与气体分压成正比。动脉血氧分压在13.3kPa (1000mmHg)时,每100ml血液中只溶解0.3mlO2,约占血液运输O2总量的1.5%。临床高压氧疗的原理,就是提高肺泡气中氧分压,使溶解于血液中的O2量增加,达到缓解缺氧的目的。
(2)化学结合是指O2和红细胞内血红蛋白(Hb)中Fe2+结合,形成氧合血红蛋白(HbO2)。因HbO2中的Fe2+仍保持低铁状态,故不是氧化作用而称氧合。正常成人每100ml动脉血中HbO2结合的O2约为19.5ml,约占血液运输O2总量的98.5%。血红蛋白与O2的结合是可逆性的,而且反应迅速,不需酶催化,主要取决于血液中氧分压。当血液流经肺部时,由于氧分压高,Hb与O2迅速结合成HbO2;而被输送到组织时,由于组织处氧分压低,HbO2则迅速解离,释放出O2,成为去氧血红蛋白。
2.二氧化碳的运输
(1)物理溶解:正常成人每100ml静脉血中CO2含量约为53%,物理溶解CO2仅3ml,约占6%。

二氧化碳的运输
(2)化学结合:CO2的化学结合运输形式有两种:
①碳酸氢盐形式:CO2以碳酸氢盐(主要是钠盐)形式运输,约占CO2运输总量的 87%,其运输过程参看下图。
当血液流经组织时,CO2顺分压差由组织扩散入血浆,因血浆中缺乏碳酸酐酶,故血浆中二氧化碳很快透入红细胞内,在碳酸酐酶的催化下,CO2迅速与H2O结合成 H2CO3,并可解离成 H+和HCO3-。由于红细胞膜对负离子容易通透,于是HCO3-除一小部分在红细胞内形成KHCO3外,大部分顺浓度差透入血浆,与血浆中Na+结合而形成NaHCO3。因正离子不能随HCO3-透出,造成红细胞膜内外电位差而影响HCO3-的透出;这时吸引Cl-向红细胞内转移,维持了膜内外电平衡,又促进了HCO3-透出,这一现象称为氯转移。可见,红细胞中碳酸酐酶的作用及氯转移的效应,使血液运输CO2能力大大增强。当静脉血流经肺部毛细血管时,CO2顺分压不断由血浆扩散入肺泡,于是上述反应向相反方向进行,CO2被释放排出。
②氨基甲酸血红蛋白形式:当血液流经组织时,进入红细胞的CO2除大部分形成HCO3-外,同时还有一部分CO2直接与血红蛋白的自由氨基结合,形成氨基甲酸血红蛋白(Hb NHCOOH),又称碳酸血红蛋白(HbCO2)。它约占CO2运输总量的7%。Hb.CO2形成后随静脉血流经肺部时,又解离释放出CO2,故HbCO3的形成和解离也是可逆的,不需酶参与。此反应进行很快,主要取决于血液中二氧化碳分压。
(七)肺容量与肺通气量
肺容量和肺通气量是衡量肺通气功能的指标,在不同状态下气量有所不同。
1.肺容量
肺容量指肺容纳的气量。在呼吸周期中,肺容量随着进出肺的气体量的变化而变化。其变化幅度主要与呼吸深度有关,可用肺量计测定和描记(见下图)。

肺容量变化的记录曲线
(1)潮气量:每次呼吸时吸入或呼出的气量,称为潮气量。正常成人平静呼吸时,潮气量为400-600ml。深呼吸时,潮气量增加。每次平静呼气终点都稳定在同一个水平上,这一水平的连线称为平静呼气基线。
(2)补吸气量和深吸气量:平静吸气末再尽力吸气所能增加的吸入气量,称为补吸气量,正常成人为1500ml-2000ml。补吸气量与潮气量之和,称为深吸气量。深吸气量是衡量肺通气潜力的一个重要指标。
(3)补呼气量:平静呼气末再尽力呼气所能增加的呼出气量,称为补呼气量。正常成人为900ml-1200ml。最大呼气终点构成了最大呼气水平。
(4)残气量和功能残气量:最大呼气末肺内残余的气量,称为残气量或余气量,正常成人为1000ml-1500ml。平静呼气未肺内存留的气量,称为功能残气量,它是补呼气量和残气量之和,正常时很稳定。肺气肿患者的功能残气量增加,呼气基线上移;肺实质性病变时则减少,呼气基线下移。
(5)肺活量和用力呼气量:最大吸气后做全力呼气,所能呼出的气量,称为肺活量,它等于深吸气量和补呼气量之和,正常成年男性约为3500ml,女性约为2500ml。
(6)肺总量:肺所能容纳的最大气量,称为肺总量,它等于肺活量与残气量之和,正常成年男性为5000ml-6000ml,女性为3500ml-4500ml。
2.肺通气量
(1)每分肺通气量:指每分钟进肺或出肺的气体总量,简称为每分通气量。其值等于潮气量与呼吸频率的乘积。正常成人安静时呼吸频率为12-18次/min,故每分通气量为6-8L。
(2)每分肺泡通气量:指每分钟进肺泡或出肺泡的有效通气量,简称为肺泡通气量。气体进出肺泡必经呼吸道,呼吸道内气体不能与血液进行气体交换,故将呼吸道称为解剖无效腔,正常成人其容积约为150ml。每次进或出肺泡的有效通气量等于潮气量和去无效腔气量,故每分肺泡通气量的计算公式如下:
每分肺泡通气量(L/min)=(潮气量一无效腔气量)×呼吸频率
正常成人安静时每分肺泡通气量约为4.2L,相当于每分通气量的70%左右。潮气量和呼吸频率的变化,对每分通气量和肺泡通气量的影响是不同的。
(六)心脏的内分泌功能
自1984年以来,国内外学者从大鼠和人的心房组织中提取纯化的心房钠尿肽以后,人们对心脏功能有了新的认识,即心脏不仅是血液循环的动力器官,而且还是一个内分泌器官。
心房钠尿肽是由心房肌细胞产生和分泌的一类具有活性的多肽,又称为心房肽或心钠素。它的主要作用是:利钠、利尿,舒张心管、降低血压。因而心钠素参与水盐平衡、体液容量和血压的调节。
(五)心脏的生理特性
心肌的生理特性包括自动节律性、传导性、兴奋性和收缩性。前三者为电生理特性,后者为机械特性。
1.自动节律性
心脏在离体和脱离神经支配下,仍能自动地产生节律性兴奋和收缩的特性,称为自动节律性(简称自律性)。心脏的自律性源于心肌自律细胞4期自动去极化。由于心脏特殊传导系统各部分的自律细胞4期去极化速度快慢不一,因而各部分的自律性高低不同。窦房结的自律性最高,约为100次/min;房室交界次之,约为50次/min;心室内传导组织最低,约为20-40次/min。正常心脏的节律活动是受自律性最高的窦房结所控制,因而窦房结是心脏兴奋的正常起搏点。其他特殊传导组织,其自律性不能表现出来,称为潜在起搏点。当窦房结的自律性异常低下,或潜在起搏点的自律性过高时,潜在起搏点的自律性就可表现出来,成为异位起搏点。以窦房结为起搏点的心脏活动,称为窦性节律;由异位起搏点引起的心脏活动,称为异位节律。
2.传导性
心脏特殊传导系统和心肌工作细胞都有传导兴奋的能力。其传导兴奋的基本原理和神经纤维相同。
正常心脏内兴奋的传导主要依靠特殊传导系统来完成。当窦房结发出兴奋后,通过心房肌传布到整个右心房和左心房;同时,沿着心房肌组成的“优势传导通路”迅速传到房室交界。心房内传导历时约0.06-0.11s。房室交界是正常兴奋由心房传入心室的惟一通路,但其传导速度缓慢,尤以结区最慢,因而占时较长,约需0.1s,这种现象称为房室延搁。房室延搁具有重要的生理意义,它使心房与心室的收缩不在同一时间进行,只有当心房兴奋和收缩完毕后才引起心室兴奋和收缩,使心室得以充分充盈血液,有利于射血。心室内特殊传导组织的传导速度快,历时约0.06-0.1s,其中以浦肯野纤维最快,只要兴奋传到浦肯野纤维,几乎立刻传到左、右心室肌,引起两心室兴奋。兴奋从窦房结传到心室肌,总共约需0.22s。现将兴奋的传导途径和速度简示如图。

传导系统任何部位发生功能障碍,都会引起传导阻滞,导致心律失常。
(四)心脏泵血过程
心脏泵血过程,左心和右心的活动基本一致。现以左心为例来讨论心脏泵血过程和泵血过程中的各种变化。
1.室缩期
根据心室内压力和容积等变化,室缩期可分为等容收缩期和射血期。
(1)等容收缩期:心室收缩前,室内压低于主动脉压和心房内压,此时半月瓣关闭而房室瓣开放,血液不断流入心室。心室收缩开始后,室内压迅速升高,在室内压超过房内压时,心室内血液推动房室瓣使其关闭,防止血液倒流入心房。但在心室内压力未超过主动脉压之前,半月瓣仍处于关闭状态。在此段时间内,由于房室瓣与半月瓣均处于关闭状态,心室容积不变,故称为等客收缩期。等容收缩期历时约0.06s,该期的长短与心肌收缩力的强弱及动脉血压的高低有关,在心肌收缩力减弱或动脉血压升高时,等容收缩期将延长。
心室收缩期 心室舒张期
心脏的射血与充盈
(2)射血期:随着心室肌的继续收缩,心室内压继续上升,一旦心室内压超过主动脉压,心室的血液将半月瓣冲开,迅速射入动脉,心室容积随之相应缩小。此期称为射血期,历时约0.24s。血液射入动脉起初,速度快,血量多,约占射血量的80%-85%;随后心室肌收缩力减弱,室内压开始下降,射血速度减慢,射血量减少。
2.室舒期
室舒期按心室内压和容积的变化可分为等容舒张期和充盈期。
(1)等容舒张期:心室舒张开始,室内压下降,在心室内压低于主动脉压时,动脉内血液逆流推动半月瓣关闭,防止血液返流入心室。当在心室内压还高于心房内压之前,房室瓣仍处于关闭状态,无血液进出心室,此时心室舒张但其容积不变,称为等容舒张期,历时约0.08s。
(2)充盈期:随着心室继续费张,室内压继续下降,直至心室内压低于房内压时,心房内的血液冲开房室瓣而流入心室,心室容积随之增大。血液不断由静脉和心房进入心室的这段时间,称为充盈期。此期历时约0.42s。起初血流速度较快、血流量较多,以后随着心室内血液增多,流入的速度减慢。在下一个心动周期的心房收缩时,血液又较快进入心室。从心室的充盈量来看,通过心房收缩进入心室的血液量仅占30%,其余大部分血液都是由于心室舒张,室内压低于房内压将血液抽吸入心室的。
总的说来,心脏泵血能按一定方向流动是取决于心瓣膜的开闭,而心瓣膜开闭又取决于心瓣膜两侧压力大小;心内压大小取决于心肌的舒缩,特别是心室肌的舒缩活动。现将心动周期中心脏内各种变化归纳如下表。
心动周期中心腔内压力、容积、辩膜、血流等变化

(三)血浆渗透压
1.血浆渗透压形成及数值
血浆渗透压由两部分溶质所形成:一部分是血浆中的无机盐、葡萄糖、尿素等小分子晶体物质形成的血浆晶体渗透压,另一部分是血浆蛋白等大分子物质所形成的血浆胶体渗透压。由于血浆中小分子晶体物质的颗粒非常多,因此血浆渗透压主要是晶体渗透压。5%葡萄糖液或0.9%NaCl溶液的渗透压与血浆渗透压相近,故称为等渗溶液。血浆胶体渗透压很小,仅为3.33kPa(25mmHg)左右。通常血浆蛋白中白蛋白的含量较多、分子量较小,因此在维持血浆胶体渗透压中,白蛋白尤为重要。
2.血浆渗透压的生理意义
血浆渗透压具有吸取水分透过生物半透膜的力量。由于细胞膜和毛细血管壁不同,因而晶体渗透压与胶体渗透压表现出不同的生理作用(如下图所示)。

血浆渗透压示意图
图示血浆与红细胞内晶体渗透压相等,可维持红细胞内外的水平衡,维持红细胞正常形态;血浆与组织液胶体渗透压不等,使组织液的水转移到血管内,维持血容量。
(1)晶体渗透压的作用:细胞膜允许水分子通过,不允许蛋白质通过,对一些无机离子如Na+、Ca2+等大多严格控制,不易过。这就造成细胞膜两侧溶液的渗透压梯度,从而导致渗透现象的产生。由于晶体比胶体溶质颗粒多,形成的渗透压高,因此血浆晶体渗透压对维持细胞内、外水分的正常交换和分布,保持红细胞的正常形态有重要作用。例如,当血浆晶体渗透压降低时,进入细胞内的水分增多,致使红细胞膨胀,直至膜破裂。红细胞破裂使血红蛋白逸出,这种现象称为溶血。反之,当血浆晶体渗透压高时,红细胞中水分渗出,从而发生皱缩。
(2)胶体渗透压的作用:毛细血管壁只允许水分子和晶体物质通过,不允许蛋白质通过,因而毛细血管内、外水分的交流取决于胶体渗透压。血浆中胶体渗透压比组织液中胶体渗透压大,故血浆胶体渗透压对调节毛细血管内外水分的正常分布,促使组织 中水分渗入毛细血管以维持血容量,具有重要作用。当血浆蛋白减少,血浆胶体渗透压降低时,组织液增多,引起水肿。
(二)肌细胞的收缩功能
人体各种形式的运动,主要是靠肌肉细胞的收缩活动来完成。不同肌肉组织在结构和功能上虽各有特点,但收缩的基本形式和原理是相似的。现以骨骼肌为例来说明。
1.骨骼肌收缩的形式
肌肉兴奋后引起的收缩,可因不同情况而有不同的收缩形式。
(1)等长收缩和等张收缩:肌肉收缩按其长度和张力的变化可分为两种:一种是肌肉收缩时,只有张力增加而长度不变的收缩,称为等长收缩;一种是肌肉收缩时,只有长度缩短而张力不变的收缩,称为等张收缩。等长收缩和等张收缩与其负荷大小有关。
肌肉承受的负荷分为前负荷和后负荷两种。前负荷是指肌肉收缩前就加在肌肉上的负荷,它可以增加肌肉收缩的初长度(收缩前的长度),进而增强肌肉收缩力。后负荷是指肌肉收缩开始时才遇到的负荷,它能阻碍肌肉的缩短。在有后负荷的情况下,肌肉不能立即缩短而增强张力,出现等长收缩。当张力增强到超过后负荷时,肌肉缩短而张力不再增加,呈现等张收缩。
在整体内,骨骼肌收缩时,既改变长度又增加张力,属于混合形式。但由于机体内肌肉的功能特点和附着部位的不同,在收缩形式上有所侧重,如咬肌收缩偏于等长收缩;眼外肌收缩偏于等张收缩。
(2)单收缩和强直收缩:整块骨骼肌或单个肌细胞受到一次短促而有效的刺激时,被刺激的肌细胞出现一次收缩过程(包括肌细胞的缩短和舒张),称为单收缩。肌肉受到连续的有效刺激时,出现强而持久的收缩,称为强直收缩。由于刺激频率不同,强直收缩又可分为不完全强直和完全强直两种。前者是新刺激引起的收缩落在前一个收缩过程的舒张期所形成的;后者是新刺激引起的收缩落在前一个收缩过程的缩短期所形成的。强直收缩是各次单收缩的融合(如下图所示),其收缩幅度比单收缩大。在整体内的各种骨骼肌收编是以整块肌肉为单位进行的。由于支配骨骼肌的运动神经冲动是连续多个动作电位,因此体内骨骼肌收缩都是强直收缩。

单收缩与强直收缩曲线图
1.收缩曲线 2.刺激记号
2.骨骼肌收缩的原理
肌细胞收缩的原理,目前用滑行学说来解释。该学说认为,肌肉的收缩并非是肌细胞(肌纤维)中构成肌原纤维的肌丝本身长度的缩短或卷曲,而是由于细肌丝向粗肌丝之间滑行,使肌节长度(两Z线间距)缩短,从而出现肌纤维和肌肉的收缩(如下图所示)。

肌丝滑行示意图
简单地说,肌肉的收缩和舒张,就是由于细肌丝向粗肌丝间滑行造成的。肌丝的滑行过程,就是在一定的Ca2+浓度下,构成细肌丝的肌动蛋白和构成粗肌丝的肌球蛋白结合和解离的过程。
(一)细胞的生物电现象
生物电现象是指生物细胞膜在安静状态和活动时伴有的电现象。它与细胞兴奋的产生和传导有着密切的关系。现以神经细胞为例来讨论细胞的生物电现象。
1.静息电位及产生原理
(1)静息电位:细胞膜处于安静状态下,存在于膜内外两侧的电位差,称为静息电位。如下图所示,将两个电极置于安静状态下神经纤维表面任何两点时,示波器屏幕上的光点在等电位线作横向扫描,表示细胞膜表面不存在电位差。但如将两个电极中的一个微电极(直径不足1μm)的尖端刺入膜内,此时示波器屏幕上光点迅速从等电位下降到一定水平继续作横向扫描,显示膜内电位比膜外电位低,表示细胞膜的内外两侧存在着跨膜电位差。此电位差即是静息电位。一般将细胞膜外电位看作零,细胞膜内电位用负值表示。
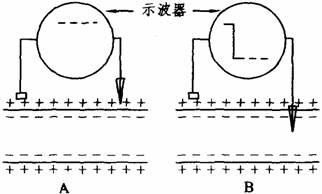
静息电位测量示意图
A.膜表面无电位差 B.膜内外两侧有电位差
同类细胞的静息电位较恒定,如哺乳类动物神经细胞的静息电位为-70--90mV。安静时,细胞膜两侧这种数值比较稳定的内负外正的状态,称为极化。极化与静息电位都是细胞处于静息状态的标志。以静息电位为准,若膜内电位向负值增大的方向变化,称为超极化;若膜内电位向负值减小的方向变化,称为去极化;细胞发生去极化后向原先的极化方向恢复,称为复极化。从生物电来看,细胞的兴奋和抑制都是以极化为基础,细胞去极化时表现为兴奋,超极化时则表现为抑制。
(2)静息电位的产生原理:“离子流学说”认为,生物电产生的前提是细胞膜内外的离子分布和浓度不同,以及在不同生理状态下,细胞膜对各种离子的通透性有差异。据测定,在静息状态下细胞膜内外主要离子分布及膜对离子的通透性见下表。
|
主要离子 |
离子浓度(mmol/L) |
膜内与膜外离子比例 |
膜对离子通透性 |
|
|
膜内 |
膜外 |
|||
|
Na+ K+ Cl- A-(蛋白质) |
14 155 8 60 |
142 5 110 15 |
1︰10 31︰1 1︰14 4︰l |
通透性很小 通透性大 通透性次之 无通透性 |
在静息状态下,由于膜内外K+存在浓度差和膜对K+有较大的通透性,因而一部分K+顺浓度差向膜外扩散,增加了膜外正电荷;虽然膜内带负电的蛋白质(A-)有随K+外流的倾向,但因膜对A-没有通透性,被阻隔在膜的内侧面。随着K+不断外流,膜外的正电荷逐渐增多,于是膜外电位上升,膜内因负电荷增多而电位下降,这样便使紧靠膜的两侧出现一个外正内负的电位差。这种电位差的存在,使K+的继续外流受到膜外正电场的排斥和膜内负电场的吸引,以致限制了K+的外流。随着电位差的增大,K+外流的阻力也随之增大。最后,当促使K+外流的浓度差和阻止K+外流的电位差所构成的两种互相抬抗的力量相等时,K+的净外流量为0,此时跨膜电位就相当于K+的平衡电位。K+的平衡电位与实际测得的静息电位略有差别,通常比测定值略高(即值略小),这是由于在静息状态下,膜对Na+也有较小的通透性,有少量Na+顺浓度差向膜内扩散的缘故。简言之,静息电位主要是K+外流所形成的电一化学平衡电位。
2.动作电位及其产生原理
 (1)动作电位:细胞膜受到刺激时,在静息电位的基础上发生一次可扩布的电位变化,称为动作电位。动作电位可用上述微电极插入细胞内测量记录下来。在测出静息电位的基础上,给予神经纤维一个有效刺激,此时在示波器屏幕上即显示出一个动作电位(如右图所示)。动作电位包括一个上升相和一个下降相,上升相表示膜的去极化过程,此时膜内原有的负电位迅速消失,并进而变为正电位,即由-70--90mV变为+20-+40mV,出现膜两侧电位倒转(外负内正),整个膜电位变化的幅度可达90-130mV。其超出零电位的部分称为超射。下降相代表膜的复极化过程,是膜内电位从上升相顶端下降到静息电位水平的过程。神经纤维的动作电位,主要部分由于幅度大、时程短(不到2ms),电位波形呈尖峰形,称为峰电位(Spike
potential)。在峰电位完全恢复到静息电位水平之前,膜两侧还有微小的连续缓慢的电变化,称为后电位。从细胞的生物电角度来看,动作电位与兴奋两者是同义语,而兴奋性是指细胞或组织产生动作电位的能力。动作电位一旦产生,细胞的兴奋性也相应发生一系列改变。从时程上来说,峰电位相当于细胞的绝对不应期;后电位的前段相当于相对不应期和超常期;后电位的后段相当于低常期(如下图所示)。膜电位恢复到静息电位水平,兴奋性也就恢复正常。
(1)动作电位:细胞膜受到刺激时,在静息电位的基础上发生一次可扩布的电位变化,称为动作电位。动作电位可用上述微电极插入细胞内测量记录下来。在测出静息电位的基础上,给予神经纤维一个有效刺激,此时在示波器屏幕上即显示出一个动作电位(如右图所示)。动作电位包括一个上升相和一个下降相,上升相表示膜的去极化过程,此时膜内原有的负电位迅速消失,并进而变为正电位,即由-70--90mV变为+20-+40mV,出现膜两侧电位倒转(外负内正),整个膜电位变化的幅度可达90-130mV。其超出零电位的部分称为超射。下降相代表膜的复极化过程,是膜内电位从上升相顶端下降到静息电位水平的过程。神经纤维的动作电位,主要部分由于幅度大、时程短(不到2ms),电位波形呈尖峰形,称为峰电位(Spike
potential)。在峰电位完全恢复到静息电位水平之前,膜两侧还有微小的连续缓慢的电变化,称为后电位。从细胞的生物电角度来看,动作电位与兴奋两者是同义语,而兴奋性是指细胞或组织产生动作电位的能力。动作电位一旦产生,细胞的兴奋性也相应发生一系列改变。从时程上来说,峰电位相当于细胞的绝对不应期;后电位的前段相当于相对不应期和超常期;后电位的后段相当于低常期(如下图所示)。膜电位恢复到静息电位水平,兴奋性也就恢复正常。
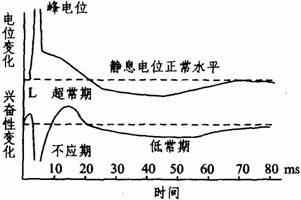
(2)动作电位的引起及产生原理:细胞膜受到刺激后,首先是该部位细胞膜上Na+通道少量开放,膜对Na+的通透性稍有增加,少量Na+由膜外流入膜内,使膜内外电位差减小,称为局部去极化或局部电位,局部电位不能远传。但Na+内流使膜内负电位减小到某一临界数值时,受刺激部位的膜上Na+通道全部开放,使膜对Na+的通透性突然增大,于是膜外Na+顺浓度差和电位差迅速大量内流,从而爆发动作电位。Na+内流是一个正反馈过程(再生性)。使膜对Na+通透性突然增大的临界膜电位数值,称为阈电位。阈电位比静息电位约小10-20mV。任何刺激必须使内负电位降到阈电位水平,才能爆发动作电位。
动作电位上升相是由于膜外Na+大量内流,膜内电位迅速高,使原来的负电位消失并高出膜外电位,在膜的两侧形成一个正外负的电位差。这种电位差的存在,使Na+的继续内流受到内正电荷的排斥,当促使Na+内流的浓度差与阻止Na+内流的位差所构成的两种互相拮抗的力量相等时,Na+的净内流停止。此时膜电位为Na+的平衡电位。简言之,动作电位的上升相是Na+内流所形成的电一化学平衡电位,是膜由K+平衡电位转为Na+平衡电位的过程。
在上升相到达Na+平衡电位时,膜上Na+通道已关闭,Na+的通透性迅速下降。与此同时,膜对K+的通透性大增。于是,K+顺浓度差和顺电位差迅速外流,使膜内外电位又恢复到原来的内负外正的静息水平,形成动作电位的下降相。简言之,动作电位下降相是K+外流所形成,是膜由Na+平衡电位转变为K+平衡电位的过程。
细胞膜在复极化后,跨膜电位虽然恢复,但膜内Na+有所增多,而K+有所减少。这时便激活了细胞膜上的钠一钾泵,通过Na+、K+的主动转运,重新将它们调整到原来静息时的水平,以维持细胞正常的兴奋性。
(3)动作电位的特点:动作电位具有“全或无”现象,刺激达不到阈强度,不能产生动作电位(无),一旦产生,幅度就达到最大值(全)。幅度不随刺激的强度增加而增加。
(4)动作电位的传导特点:动作电位在同一细胞沿膜由近及远地扩布称为动作电位的传导。其传导特点有:①不衰减性传导。动作电位传导时,电位幅度不会因距离增大而减小。②双向性传导。如果刺激神经纤维中段,产生的动作电位可从产生部位沿膜向两端传导。
湖北省互联网违法和不良信息举报平台 | 网上有害信息举报专区 | 电信诈骗举报专区 | 涉历史虚无主义有害信息举报专区 | 涉企侵权举报专区
违法和不良信息举报电话:027-86699610 举报邮箱:58377363@163.com