题目列表(包括答案和解析)
根据最新国际生物学奥林匹克竞赛(IBO)纲要和全国中学生生物学竞赛大纲(试行)要求,有关植物生理学的内容主要包括:水分、矿质营养的吸收和运输;光合作用;呼吸作用;蒸腾作用;生长和发育,激素;生殖。上述内容中,原中学生物教学大纲中已有的不再重复,只对其他一些在竞赛中经常遇到但又难于理解的内容作些简要的分析或说明。
例1 根据遗传学的有关知识,分析回答下列问题:
(1)噬菌体фX174是单链DNA生物,当它感染宿主细胞时,首先形成复制型(RF)的双链DNA分子。如果该生物DNA的碱基构成是:0.27A,0.31G,0.22T和0.20C。那么,RF及与亲链互补的DNA链的碱基构成将会怎样?
(2)若某高等生物的蛋白质由600个氨基酸组成,那么,为该蛋白质编码的基因至少有多长(单位:米)?
分析 (1l)根据DNA双链形成中的碱基互补规律推知,与亲链互补的单链DNA,其碱基构成应与亲链DNA中与之互补的碱基含量对应相等,即0.22A,0.20G,0.27T和0.31C。而RF的碱某构成应是A=T=E=(0.27+0.22)/ 2=0.245;C=G=(0.31+0.20)/ 2=0.255。
(2)因为每一种氨基酸的密码子为mRNA上的三个相邻碱基,而mRNA上三个相邻碱基还有可能成为终止密码,即不编码对应的氨基酸。因此为600个氨基酸编码的mRNA碱基数目至少为600×3=1800个,那么转录该mRNA的基因至少有1800个碱基对。根据DNA双螺旋结构模型,邻近碱基对之间的距离为3.4Å,1 Å=10-10m,则该基因的长度至少为:
1800×3.4×10-10=6.12×10-7(米)
(九)人类的起源与进化
南猿阶段→能人阶段→直立人阶段→智人阶段。
1.从猿到人的过渡阶段(南方古猿):从1924年首先发现南方古猿化石(南非汤恩),1973年在坦桑尼亚莱托利和埃塞俄比亚哈达发现了阿法南猿(南方古猿阿法种)的化石,命名为“露西”,距今约300万至360万年前,这是目前发现的最古老的人类化石。
2.早期猿人(能人):1959年-1960年在坦桑尼亚奥杜韦找到能人化石,是这一阶段的代表,生活于距今约160万至300万年前。能人是否为真正人类尚未有定论。
3.晚期猿人(直立人):第一个人属化石是荷兰人杜布瓦于1890年-1891年在印尼发现的爪哇直立人。直立人已是真正的人类了,可能有一定的语言能力,如北京猿人、蓝田猿人、元谋猿人、德国海德堡猿人等。距今约30万至180万年前。
4.智人:①早期智人(古人)。出现最早的智人是尼安德特人(尼人),1856年在德国发现的。我国发现的早期智人有陕西大荔人、广东马坝人、湖北长阳人、山西丁村人等。生活于距今10万至30万年前。②晚期智人(新人)。最早发现的新人化石是法国克罗马农人。我国境内发现的晚期智人中最著名的是周口店的“山顶洞人”(一般认为是原始黄种人)和广西的柳江人。晚期智人生活于距今约3万至4万年前。
(八)物种形成的两种方式:渐进式与爆发式
物种形成是一个由量变到质变的过程。主要有两种不同的方式:渐进式和爆发式。
1.渐进式的物种形成
又可分为两种方式,即继承式的物种起源和分化式的物种形成。继承式的物种形成是指一个在同一地区内逐渐演变成另一个种。这种进化方式,由于所要时间很长,所以无法见到。当演变成另一个新种后,原来的物种便逐渐被自然选择所淘汰,不复存在。例如马的进化,原始马有五个脚趾,现代马只有一个脚趾,现在再也找不到多趾马了。
分化式物种形成是指一个物种在其分布范围内逐渐分化成两个以上的物种。地理隔离是导致这类物种形成方式的基本因素。达尔文在加拉帕戈斯群岛所发现的许多种鸡禽就是其中一例。这些鸣禽的祖先是由于偶然的原因从南美大陆迁来的,它们逐渐分布到各个岛上,由于水域相隔又由于每个岛上的生物类型是从邻近大陆上迁移过来的,同时又各自产生了适应于当地环境的新的类型。总之,渐变式的物种形成主要是通过变异的逐渐积累而形成亚种,再由亚种形成一个或多个新种。
2.爆发式物种形成
这是一种比较迅速的物种形成方式。在植物界,通过种间或属间远缘杂交和杂种的染色体加倍,可以在较短时间内形成新的物种,异源多倍体的形成就属这种方式。例如根据小麦种、属间大量远缘杂交研究分析,证明普通小麦起源于两个不同的亲缘属,逐步地通过属间杂交和染色体加倍,形成了异源六倍体普通小麦。
(七)多分子体系的形成与原始生命的出现
生物大分子还不是原始生命,蛋白质及核酸都是比较容易被破坏的大分子,它们在海水中存在,在较高的温度下,都会被破坏。如果要使它们存在下来,除了环境条件合适以外,它们必须要结合起来形成多分子体系。并且这些生物大分子在单独存在的情况下也不表现出生命的现象,只有当它们形成多分子体系时,才显示出一些生命现象。这种多分子体系就是原始生命的萌芽。
多分子体系是如何形成的?第一步还是浓缩。生物大分子形成之后,在海水中继续由于蒸发作用而浓缩;它们也可以通过吸附于粘土表面而浓缩在一起。浓缩引起了第二步改变,这第二步改变目前有些不同的看法,主要是团聚体学说及微球体学说。
团聚体乃是胶粒的凝聚作用形成的。奥巴林首先提出了生物大分子形成团聚体乃是生命发展过程中一个主要阶段。他用实验方法获得了团聚体小滴,发现它有一定的生命现象。
奥巴林做了一系列试验,用白明胶的水溶液与阿拉伯胶的水溶液混在一起,混合之前,这两种溶液都是透明的,混合之后就变混浊了。在显微镜下观察就可以看到在均匀的溶液中出现了小滴,即团聚体。它们四周与溶液的水有明显的界限。用蛋白质、核酸、多糖、磷脂、多肽及多孩青酸等溶液也能形成这样的团聚体。
团聚体表现出一定的生命现象。例如,团聚体能从周围的水溶液中吸取各种不同的物质,这样,团聚体就可以长大,好像生长一样。长到一定程度时,团聚体还能“出芽”,形成小的团聚体,如同生殖一样。团聚体吸收外界物质似乎也有选择性。团聚体形成后内部具有一定的结构。团聚体如吸收了酶系,酶可以在团聚体内进行工作,如合成或分解某些物质,使团聚体相应地增加或减少某些物质。团聚体与周围环境有一个明显的界限,这是原始界膜形成的一种可能方式。
这许多特征使人想像,多核苷酸溶液与多肽溶液,或核酸溶液与蛋白质溶液,在浓缩后,在一定的温度与酸度的条件下,形成了团聚体这一多分子体系;这是原始生命形成过程中一个重要阶段。有人曾在数百米至数千米海水深处,发现有类似团聚体的东西,这是一个直接证明。
另一个学说就是微球体学说。这一学说认为类蛋白体形成的微球体乃是最初的多分子体系。把一个热的多肽溶液冷却时就会形成许多小球,这些小球也表现出很多生物学特性。例如,它们能吸收外界物质,也能“出芽”生殖,它们在高渗溶液中收缩。在低渗溶液中膨胀,它们具有一个双层膜,内部具有一定结构,并且表现出类似细胞质流动的活动(当它吸收了腺三磷之后)。最后,它们可以聚集起来,像群集在一起的细菌一样。
这两个学说究竟哪一个正确有待进一步研究,但是不论是哪一种多分子体系,有三点是同样重要的。
第一,就是这个多分子体系内部具有一定的物理化学结构,这是生命起源的一个主要条件。有了一定的物理化学结构后,才有吸收物质及进行其他化学反应的能力,并且这些反应都能以一定方式进行。分子在空间的有规则的排列是造成这些特性的主要根据。同时这样的多分子体系才能完全脱离外界环境的影响而独立“生活”,也即有了结构就不容易被破坏。
第二,就是这个多分子体系必须同时具有蛋白质及核酸。假如是团聚体,这个团聚体必须是由核酸溶液与蛋白质溶液,或多肽溶液与多孩音酸溶液所组成。假如是微球体,那么它必须吸收入核酸或多核苷酸进入它的组成。可以想像,在早期团聚体有多种多样的,例如,白明胶溶液与阿拉伯胶溶液形成的团聚体。微球体也可能只是类蛋白球小体,其中没有核酸。它们都在自然选择中被淘汰了。只有同时具有核酸及蛋白质的多分子体系才被选择保留下来。因为单有一种是不够的,核酸不具有酶的催化作用,蛋白质一般不具有复制性能,只有二者配合在一起,才能形成一个完整的调节系统。至于核苷酸如何与氨基酸形成相应的关系(核酸密码),现在还不知道,可能也是长时期的偶然配合及选择的结果。总之,蛋白质与核酸结合在一起的多分子体系才是真正原始生命,因为生命的物质基础是核酸蛋白。其他大分子结合成的多分子体系不能发展成为原始的生命,但是其他的大分子及分子,如多糖、脂肪等也可以被吸收入原始生命的多分子体系中。
这里可以看出,自然选择远在真正生命起始之前已经开始了作用。各种多分子体系是经过自然选择之后才形成具有生命性能的多分子体系的。
第三,就是原始膜的形成。在原始生命的出现和演化为以后的细胞形态的生命过程中,膜的发生和发展是极为重要的;它使多分子体系与原始海洋隔离开来,形成一个独立的体系。只有内部一定的结构而没有原始膜是不能形成独立体系的。
原始膜生成之后,多分子体系就形成了一个独立系统,通过原始膜的控制,与周围环境发生物质交换。原始膜控制了某些物质的能进入与不能进入,也就控制了环境对于生物系统的作用。随着原始生命的不断进化,结构、功能不断完善和复杂化,逐步形成了现在的生物膜。
(六)基因频率和哈代--温伯格平衡
群体中的基因频率是指某一等位基因在所有等位基因总数中所出现的百分率。例如,在人类的MN血型系统中,基因型LMLM的个体表现为M血型;基因型LMLN的个体表现为MN血型;基因型LNLN的个体表现为N血型。1977年,上海中心血防站调查了1788个MN血型者,发现有397人是M血型,861人是MN血型,530人是N血型。假如用基因频率表示,其结论是:
LM基因的频率p=(2×397+861)/(2×1788)=0.4628
LN基因的频率q=(861+2×530)/(2×1788)=0.5372
群体中的基因频率能否保持稳定?在什么情况下才能保持稳定?对此,英国数学家哈代和德国医生温伯格分别于1908年和1909年提出一个理论,即如果有一个群体凡符合下列条件:1)群体是极大的;2)群体中个体间的交配是随机的;3)没有突变产生;4)没有种群间个体的迁移或基因交流;5)没有自然选择,那么这个群体中各基因频率和基因型频率就可一代代稳定不变,保持平衡。这个理论称哈代--温伯格平衡,也称道传平衡定律。
哈代--温伯格平衡可用一对等位基因来说明。一个杂合的群体中,在许多基因位点上,可以有两个或两个以上的等位基因。但只要这个群体符合上述5个条件,那么其中杂合基因的基因频率和基因型频率,都应该保持遗传平衡。设一对等位基因为A与a,亲代为AA与aa两种基因型,其基因频率分别为p与q(因为是百分率,所以p+q1)。自由交配后,按孟德尔遗传法则确定F1代具有AA、Aa、aa3种基因型。其频率如下列公式所示:
p2AA+2pqAa+q2aa=1
假定3种基因型提供等量的配子输送给群体(基因库),其中纯合子AA与aa只产生一种配子(A或a),杂合子Aa产生两种配子(A与a),那么F2代的A与a两种配子的频率为:
A=p2+1/2×2pq=p2+pq=p(p+q)=p
a=q2+1/2×2pq=q2+pq=q(p+q)=q
结果配子的基因频率与亲代完全相同,3种基因型的频率也不变。以此类推,其后代的情况同样如此,群体保持其稳定平衡。上述情况说明,一个群体总是倾向于保持其原有的变异结构或组成。
基因在群体中所占的比例称为基因频率,而不同基因型在群体中所占的比例称为基因型频率。假定等位基因为Aa,则A与a的频率为基因频率,分别用p、q表示,AA、Aa、aa的频率为基因型频率,分别为P、H和Q,则有p+q=l,P+H+Q=1。基因频率与基因型频率的关系是:
p=P+1 / 2H,q=Q+1 / 2H,P=p2,H=2pq,Q=q2,p2+2pq+q2=1
基因频率决定了基因型的频率,但在实际计算时则基因频率是由基因型频率推算出来的,而基因型频率又是由表型频率估算出来的。
哈代--温伯格的发现说明了在一定条件下群体可以保持遗传平衡,但在事实上,这些条件基本上是不存在的。因为所谓极大的群体是不存在的,群体内个体之间充分的随机交配也不现实,突变总不断在产生,外来基因由于流动与迁移不断在加入,自然选择也时时在发生。因此,这一定律恰恰证明遗传平衡是相对的,而群体的基因频率一直在改变,进化是不可避免的。
(五)性别决定
性别,即生物个体雌雄的差别,是生物体一种特殊的性状,有人称为性性状。和其他性状一样,性别受遗传物质的控制,同时也受环境因素的影响。决定生物雌雄性别发展趋势的内在因素和方式称为性别决定。由性染色体决定性别是生物界普遍存在的一种性别决定机制。所谓性染色体是与性别决定直接相关的染色体,例如人类和果蝇的X与Y染色体,鸟类的Z和W染色体。性染色体是麦克朗(1901)在直翅目昆虫中首先发现的,在生物体细胞中通常成对存在,但是有的生物含有一个或两个以上性染色体。细胞中性染色体以外的染色体称为常染色体。由性染色体决定性别的生物,主要有以下两种类型:
1.XY型(含XO型):
即雄性含有两个异型性染色体,雌性含有两个同型性染色体的生物。这类生物的雄性个体产生分别含有X染色体或Y染色体的两种不同的雄配子,称为异配性别。雌性个体只产生一种含X染色体的雌配子,称为同配性别。含有X染色体的雄配子与雌配子结合,产生的后代个体是雌性(XX),含有Y染色体的雄配子和雌配子结合,后代则是雄性(XY)。显然,后代个体的性别是由父方决定的。由于雄性个体产生含X染色体或Y染色体的两种配子数目相等,所以生物种群的两性比例为1︰1。XY型在生物界普遍存在,例如人类、哺乳类,某些鱼类、两栖类、双翅目昆虫以及某些雌雄异林的植物都属于XY型。
XO型:是XY型中一个特殊的类群。它的雄性个体只有一条X染色体,没有Y染色体,其性染色体组成为XO,故称XO型。但是雌性有两条X染色体,其性染色体组成为XX。例如,蝗虫雌性个体细胞染色体数是:2n=24,XX,雄性是2n=23,XO。此外,蟑螂、蟋蟀、虱等也属于这种类型。
2.ZW型(含ZO型):
与XY型相反,凡雌性含有两个异型性染色体,雄性含有两个同型性染色体的生物,称为ZW型。ZW型的雌性个体为异配性别,雄性个体为同配性别,后代的性别是由母方而不是由父方决定的。鸟类、某些鱼类、两栖类、爬行类、鳞翅目昆虫等属于ZW型。
ZO型;与ZW型相类似。但是ZO型生物的雌性个体只有一个Z染色体而没有W染色体,而雄性个体有两条Z染色体。ZO型生物较少见,例如鸡、鸭等家禽,某些鱼类和鳞翅目昆虫等属于此类。
生物的性别决定机制多种多样。例如蜜蜂和蚂蚁等昆虫是由细胞的染色体倍数决定雌雄性别的。由正常受精卵发育成的二倍体(如蜂王)表现为雌性,由未受精卵发育成的单倍体(如雄蜂)则表现雄性。但是蜜蜂雌性的发育还与食物有关。二倍体幼虫食用蜂王浆的发育成蜂王,食蜂蜜和花粉的发育成工蜂一一不育的雌性。玉米的性别分化至少与两对基因有关。正常的玉米植株为雌雄同株异花,单性的雄性植株和单性的雌性植株均是由有关基因不同的基因型决定的。海生蠕虫后螠如果以自由生活方式为生表现为雌性,如果在雌虫体内营寄生生活则为雄性。
(四)连锁与互换规律
连锁遗传现象是1906年美国学者贝特森和潘耐特在香豌豆两对相对性状的杂交试验中首先发现的。他们观察到同一亲本所具有的两个性状在杂交后代中常有相伴遗传的倾向,这就是性状的连锁遗传现象。1911年美国遗传学家摩尔根根据在果蝇中发现的类似现象提出连锁和交换的理论,从而确立了遗传学第三定律--基因的连锁互换定律。
1.不完全连锁和完全连锁
(1)果蝇的性状连锁遗传:摩尔根用灰身长翅纯系果蝇与黑身残翅纯系果蝇杂交,F1都表现灰身长翅。让F1的雌蝇与黑身残翅雄蝇交配,测交后代(Ft)的表现型类型及其数目是:21灰长(42%),4灰残(8%),4黑长(8%),21黑残(4%)。
在上述测交子代群体中,亲本型(灰长和黑残)占84%,明显多于占16%的重组型(灰残和黑长)。显然F1雌蝇产生的四种类型的配子数目是不等的,即亲型配子远多于重组型配子。
(2)基因的连锁和交换:摩尔根等人用基因的连锁和交换理论解释性状连锁遗传现象,其基本要点如下:
①控制不同性状的非等位基因,位于同一对同源染色体上的不同基因座位上。同一条染色体上彼此连锁的基因群,称为一个连锁群。
②连锁基因常常联系在一起不相分离,随配子共同传递到下一代。这种位于同一染色体上的基因相伴遗传的现象称为基因连锁。
③在减数分裂中,可能发生同源染色体的非姊妹染色单体之间对应片段的交换,一旦交换发生在连锁基因之间,使位于交换片段上的等位基因互换,从而导致非等位基因间的基因重组。由于同源染色体之间发生交换,而使原来在同一染色体的基因不再伴同遗传的现象称为基因交换。连锁的基因之间能够发生交换,称为不完全连锁。例如,F1灰身长翅雌蝇的基因型为BV / bv,在形成配子时,如果在B-V之间发生交换,那么它将产生BV、bv、Bv、bV四种配子。
④如果连锁基因之间发生一次交换(单交换),该交换只涉及同源染色体的两条非姊妹染色单体,那么,F1每个发生交换的性母细胞将只会产生一半重组型配予,另一半是亲本型配子。但是交换是较少发生的事件,进行减数分裂的全部性母细胞,不可能都发生完全相同的交换重组过程。因此F1产生的配子总数中,大部分(上例中为84%)是两种亲型配子,少部分(上例中为16%)为两种重组型配子。也就是说,连锁状态的两对基因杂种产生的重组型配子总是少于亲型配子,即少于配子总数的二分之一。
基因的连锁与互换理论和前述果蝇杂交实验结果是一致的(如下图所示)。
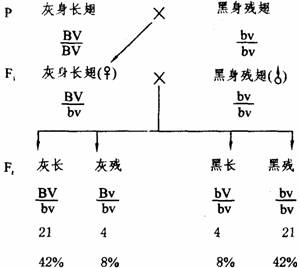
果蝇体色和翅的遗传
(3)完全连锁:
如果用上述果蝇杂交的F1灰身长翅雄蝇与黑身残翅雌蝇进行测交,其后代只有发长和黑残两种类型果蝇,而且各占一半。
F1中只有灰身长翅和黑身残翅两种亲组合,说明F1雄蝇只形成了两种亲型配子,而没有产生重组型配子。也就是说F1雄蝇在减数分裂时同源染色体之间没有发生基因交换,使连锁的基因不出现重组。这种连锁基因之间不发生交换,从而不出现基因重组的基因连锁称为完全连锁。细胞中数以万计的基因存在于为数不多的染色体上,基因连锁是必然的。同时连锁对于生命的延续也属必要,这样能够保证在细胞分裂过程中每一个子细胞都能准确地获得每一个基因。然而,完全连锁的现象是非常罕见的,迄今为止,只发现雄果蝇和雌家蚕表现完全连锁。不完全连锁基因之间发生交换已被广泛的事实证明。一般认为减数分裂时见到的同源染色体交叉现象,可以作为基因交换的细胞学证据。
2.交换值与遗传距离
交换值,通常也称为重组率,是指重组型配子占总配子数的百分率,用以表示连锁基因之间发生交换的频率大小。计算交换值的公式是:
交换值(RF)=重组型配子 / 总配子数(亲配子数十重组型配子数)×100%
测定重组型配子数最常用的方法是使F1与隐性纯合体测交,根据测交后代(Ft)中重新组合类型的数目直接确定重组型配子的数目。对于水稻、豌豆等自花授粉作物,由于杂交比较困难,可以利用F1的自交后代(F2)计算重组型配子的数目,测定其交换值。
交换值的大小变动在0-50%之间。当非等位基因是不完全连锁时,交换值总是大于0而小于50%。如果测定交换值为0,说明有关基因是完全连锁的。交换值为50%时,两个被测定的非等位基因表现自由组合。
大量的遗传学研究表明,连锁基因间的交换值是相对稳定的。交换值越小,即基因间发生交换的频率越低,表示连锁强度越大;交换值越大,则基因的连锁强度越小。一般认为基因间的连锁强度是由基因在同一染色体上的相对距离(或称遗传距离)决定的,所以通常用交换值的大小来表示连锁基因间的距离,以1%交换值作为一个距离单位(图距单位),或称厘摩(cM)。
3.基因定位与连锁图
基因定位就是确定基因在染色体上的位置,其主要内容是确定基因之间的距离和顺序。只要准确地估算出连锁基因的交换值,就能确定基因之间的遗传距离。根据紧密连锁的多个基因之间的距离,可以决定它们之间的相对顺序。将生物已知基因的相对位置标记在染色体上,绘制成图,称为连锁图或遗传学图。两点测验和三点测验是经典遗传学中基因定位的主要方法。
(1)两点测验:
两点测验又称两点测交,是基因定位最基本的一种方法。两点测验首先进行杂交获得双基因杂种(F1),然后对F1进行测交,以判断这两对基因是否连锁。如果是连锁的,根据其交换值确定它们在同一染色体上的遗传距离。前面提到的果蝇测交试验(上页图)就是一次两点测验。根据测交结果,b和v之间的交换值:
RF(b-v)=(4+4)/(21+21+4+4)×100%=16%
因此b-v之间的遗传距离为16cM(图距单位)。
 如果对紧密连锁的三个基因a、b、c分别进行三次两点测验,每两个基因之间的距离分别是:a-b为5cM,b-c为10cM,a-c为15cM,那么,连锁基因a、b、c在同一染色体上的连锁如右图。
如果对紧密连锁的三个基因a、b、c分别进行三次两点测验,每两个基因之间的距离分别是:a-b为5cM,b-c为10cM,a-c为15cM,那么,连锁基因a、b、c在同一染色体上的连锁如右图。
(2)三点测验:
根据连锁的三个非等位基因的交换行为确定它们在同一染色体上相对位置的杂交试验称为三点测验,又称三点测交。它是基因定位最常用的方法。三点测验的主要过程是:通过杂交获得三对基因杂种(F1),再使F1与三隐性基因纯合体测交,通过对测交后代(Ft)表现型及其数目的分析,分别计算三个连锁基因之间的交换值,从而确定这三个基因在同一染色体上的顺序和距离。通过一次三点测验可以同时确定三个连锁基因的位置,即相当于进行三次两点测验,而且能在试验中检测到所发生的双交换。此外,三点测验中得到的三个交换值是在相同的遗传背景和环境条件下取得的,因此使估算的交换值更加准确。现在以玉米籽粒的饱满(Sh)与凹陷(sh),非糯性(Wx)与糯性(wx),有色(C)与无色(c)三对性状的杂交为例,说明三点测验的具体步骤。为了方便起见,以“+’代表各显性基因,其对应的隐性基因仍分别以sh,wx和c表示。
三点测验的主要步骤:
(1)通过杂交和测交获得 F1的测交后代(Ft),其过程如下所示:


(2)根据F1确定连锁基因的顺序:
从上述测交后代(Ft)的资料可以看出,在群体中亲型个体①和②数目最多(2708+2538),无疑是两种亲型配子(sh++和+wx c)受精产生的。⑦和⑧两种个体数目最少(4+2),是双交换型配子受精的结果。所谓双交换型配子,是在三个连锁基因所在区域内同时发生二次交换所产生的配子。例如下图所示:

+b+和a+c就是双交换型配子。
根据两个杂交亲本的表现型推测,F1中三个连锁基因的顺序有三种可能:一是 wx在sh和c之间,即: ;二是sh在wx和c之间,即:
;二是sh在wx和c之间,即: ;三是c在sh和wx之间,即:
;三是c在sh和wx之间,即: 。这三者之中,只有第二种情况才能产生+++和sh wx c。两种双交换型配子,其他两种情况都不可能产生。据此可以确定三个连锁基因在染色体上的次序是sh位于 wx和c之间,即:。
。这三者之中,只有第二种情况才能产生+++和sh wx c。两种双交换型配子,其他两种情况都不可能产生。据此可以确定三个连锁基因在染色体上的次序是sh位于 wx和c之间,即:。
(3)计算交换值,确定基因距离:
首先分别计算wx–sh和sh–c的交换值,确定它们之间的遗传距离。
在杂交亲本产生的亲型配子中,sh与wx之间的连锁状态是sh+和+wx,即相斥相,但是F1产生的③、④、⑦、⑧四种配子中这两个基因是++和sh wx,即相引相,可见它们是上sh–wx之间发生交换形成的重组型配子。因此,sh–wx之间的交换值是:
RF(sh–wx)=(③+④+⑦+⑧)/ 总配子数×100%
=(626+601+4+2)/ 6708×100%=18.4%
同理,sh–c之间发生交换的重组型配子是⑤、⑥、⑦、⑧,那么sh–c 的交换值是:
RF(sh–wx)=(⑤+⑥+⑦+⑧)/ 总配子数×100%
=(113+116+4+2)/ 6708×100%=3.5%
根据基因在染色体上呈直线排列的原理,
 RF(wx–c)=18.4%+3.5%=21.9%
RF(wx–c)=18.4%+3.5%=21.9%
基因之间的距离分别是:wx–sh为18.4cM,sh–c为3.5cM,wx–c为21.9cM。这三个基因的连锁图表示如右图所示。
(三)孟德尔定律的扩充
孟德尔通过豌豆的杂交试验发现了基因的分离规律和自由组合规律两个定律。后来人们又用其他生物材料做实验,包括从病毒、细菌直到人,将孟德尔定律更进一步扩充。
1.不完全显性
孟德尔杂交试验所选用的生物材料,确定的几对性状,显性是完全的。然而生物界中也有其他情况。在家蚕中,黑缟对白斑是由一对等位基因决定的。这两种家蚕杂交,得到的子一代全是淡黑缟蚕。子一代雌雄个体相互交配得到的子二代中,大约是1/4黑缟、2/4淡黑缟、1/4白斑。由此看出,杂交的子一代出现了中间性状;但子二代又出现了分离,然而也有部分仍表现出中间性状。这是一种不完全显性现象,实际上也是遵循孟德尔定律的(如下图所示)。

2.嵌镶显性
嵌镶显性是我国遗传学家谈家桢教授所发现的。在异色瓢虫中,鞘翅有很多色斑变异,表现在不同的黑色斑纹上:黑线型的前缘呈黑色,均色型的后缘呈黑色。鞘翅的底色呈黄色。如果将这两种类型的纯合体交配,子一代的杂种的鞘翅出现了新的色斑,似乎是两个亲体的鞘翅重叠起来,亲代的两种黑斑都在子一代表现出来。子一代相互交配,在子二代中 1/4是黑缘型、1/4是均色型、其余一半的色斑和子一代相同(如下图所示)。嵌镶遗传现象表明;显性,不一定有隐性和它相对存在,一对性状,可以都是显性,而在生物体不同部位表现出来。
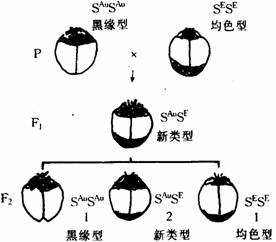
瓢虫鞘翅色斑的遗传
3.致死因子
 有一种家鼠,皮毛黄色对灰色是由一对等位基因控制的。当用黄色鼠和灰色鼠杂交,得到的子一代黄色和灰色两种鼠的比例是1︰1。将子一代中黄色鼠自相交配,得到的子二代中,黄色和灰色两种鼠的比例是2︰1。从表面上看,似乎是违反了孟德尔定律。后来研究知道,原来黄色家鼠基因是杂合体,没有纯合体的黄色家鼠。其原因是黄色家鼠基因若为纯合体是致死的,在胚胎期已经死亡而为母体吸收。所以黄色家鼠的繁殖,其后代有黄色和灰色两种,比例是2︰1(如右图所示)。
有一种家鼠,皮毛黄色对灰色是由一对等位基因控制的。当用黄色鼠和灰色鼠杂交,得到的子一代黄色和灰色两种鼠的比例是1︰1。将子一代中黄色鼠自相交配,得到的子二代中,黄色和灰色两种鼠的比例是2︰1。从表面上看,似乎是违反了孟德尔定律。后来研究知道,原来黄色家鼠基因是杂合体,没有纯合体的黄色家鼠。其原因是黄色家鼠基因若为纯合体是致死的,在胚胎期已经死亡而为母体吸收。所以黄色家鼠的繁殖,其后代有黄色和灰色两种,比例是2︰1(如右图所示)。
可见,孟德尔的分离定律中3︰1的比数是有条件的,其中之一是各基因型的生活力相等。
复等位基因:前面所涉及的都是一对等位基因,其实生物细胞中许多基因有很多等位形式,这样的基因叫复等位基因。
人类的ABO血型可作为复等位基因的例子。三个复等位基因IA、IB、i,其中IA、IB对i是显性,IA、IB为共显性。在杂合体中,一对等位基因都显示出来的现象称为共显性。三个复等位基因决定了六种基因型:IAIA、IAi、IBIB、IBi、IAIB、ii。分成A型、B型、AB型和O型4种血型。
如果血型是AB型的男人和血型是O型的女人结婚,他们子女的血型应该是A型或是B型,而不会是其他类型。从血型上看,子代不像亲代,然而却是遗传的。
ABO血型的人红细胞上有抗原(为一种糖脂),体内还有天然抗体(为蛋白质),所以输血时要特别注意。
4.抑制基国
家蚕由于品种不同,有结黄茧的,有结白茧的。白茧的又有中国品种和欧洲品种。结黄茧的和给白茧的欧洲种交配,子一代全是结白茧的;但是如果和中国种交配,子一代全是结黄茧的。这表明对于蚕的结黄茧性状来说,欧洲品种结白茧的性状是显性,中国品种结白茧的性状是隐性。把结黄茧的和欧洲结白茧的交配,得到的子一代家蚕再相互杂交,得到的子二代中,结白茧的和结黄茧的比率是13︰3。如何解释这种现象呢?
假定黄茧基因是Y,白茧基因是y,另外还有一个非等位基因I。当有I存在时,可以抑制Y的作用。这样,黄茧品种的基因型是 iiYY,欧洲白茧品种的基因型是IIyy,子一代的基因型是IiYy。因为I对Y有抑制作用,所以子一代是白茧蚕。子一代相互交配,在子二代中出现9种基因型。只要子二代个体中有I存在,Y的作用被抑制,其表型则是结白茧的。所以子二代中,结白茧的与结黄茧的比率为13(9+3+1)︰3。
一个基因抑制非等位基因的另一基因的作用,使其不能显示出来,这种基因叫做抑制基因。
5.上位效应
家兔中的灰免和白兔杂交,子一代全是灰兔。子一代灰兔相互交配;子二代中有灰兔、黑免和白兔三种兔出现,其比率为9︰3︰4。这个比率又如何解释呢?
我们发现在9︰3︰4的比率中,有色(灰+黑)︰白色是3︰1;灰色︰黑色也是3︰1。出现了两个3︰1,就可认为这里包括两对基因之差。设一对基因为C、c,另一对基因为G、g,这两对基因共同决定兔毛的颜色。当显性基因C存在时,基因型GG或Gg的表型为灰色,gg的表型为黑色,当显性基因C不存在。而只有隐性基因c时,无论是GG、Gg的基因型,还是gg的基因型,其表型都为白色。
存在有两对非等位基因,其中一对中的显性基因(或隐性基因)可遮盖另一对非等位基因的表现,这样的两对基因的关系称为上位效应。上述例子中,是隐性基因c起作用,这种现象称为隐性上位作用。若发生显性上位作用,则后代中F2会出现12︰3︰1的分离比。
(二)染色体的畸变
1.染色体结构的改变
因为一个染色体上排列着较多基因,所以不仅染色体数目的变异可以引起遗传信息的改变,而且染色体结构的变化,也可引起遗传信息的改变。
一般认为,染色体的结构变异起因于染色体或它的亚单位--染色单体的断裂。每一断裂产生两个断裂端,这些断裂端可以沿着下面三条途径中的一条发展:
(1)它们保持原状,不愈合,没有着丝粒的染色体片段最后丢失。
(2)同一断裂的两个断裂端重新愈合或重建,回复到原来的染色体结构。
(3)某一断裂的一个或两个断裂端,可以跟另一断裂所产生的断裂端连接,引起非重建性愈合。

染色体的畸变
染色体重排。每一行中上面的染色体具有正常序列的遗传信息,下面的染色体则具有重排序列。结构杂台子在减数分裂前期,正常的和重排的染色体治其长度配对,所有同源区尽可能进行联会。在缺失(a)和重复(b)的杂合子中一个染色体上有一段没有同源区配对,因此有一小段不能联会的区段。在倒位杂合子(c)中只有形成倒位环才能完全配对。在易位杂合于(d)中两个正常的和两个易位的染色体在联会中出现+字形结构。最右边的图表示果蝇结构杂合于唾液腺染色体的正常的和重排的区段
依据断裂的数目和位置,断裂端是否连接,以及连接的方式,可以产生各种染色体变异,主要的有下列四种(如上图所示)。
(1)缺失--染色体失去了片段;
(2)重复--染色体增加了片段;
(3)倒位--染色体片段作180°的颠倒,造成染色体内的重新排列;
(4)易位--非同源染色体间相互交换染色体片段,造成染色体间的重新排列。
2.染色体数目的改变
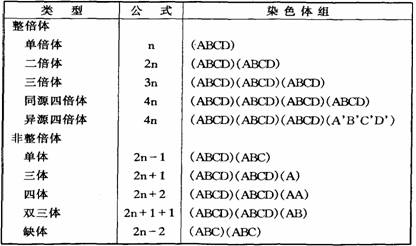
遗传学上把一个配子的染色体数,称为染色体组,用n表示。凡是细胞核中含有一个完整染色体组的,就叫做单倍体,如蜜蜂的雄蜂,n=16;含有两个染色体组的叫做二倍体,如人2n=46;有三个染色体组的叫做三倍体,如三倍体西瓜,3n=33,依此类推。这类染色体数的变化是以染色体组为单位的增减,所以称作倍数性改变,超过两个染色体组的,通称多倍体。另一类染色体数的变化是细胞核内的染色体数不是完整的倍数,通常以二倍体(2n)染色体数作为标准,在这基础上增减个别几个染色体,所以属于非整倍性改变。例如 2n-1是单体,2n-2是缺体,2n+1是三体等。
染色体的数目变异,可作以下的分类:
湖北省互联网违法和不良信息举报平台 | 网上有害信息举报专区 | 电信诈骗举报专区 | 涉历史虚无主义有害信息举报专区 | 涉企侵权举报专区
违法和不良信息举报电话:027-86699610 举报邮箱:58377363@163.com