题目列表(包括答案和解析)
(四)呼吸作用
呼吸作用为生物体进行生命活动提供能量,任何活的细胞都在不停地进行着呼吸作用,呼吸作用的停止就意味着细胞的死亡。呼吸作用分为有氧呼吸和无氧呼吸两种类型。
1.有氧呼吸的全过程
细胞有氧呼吸的全过程可分为以下三个步骤:
糖酵解:将一分子葡萄糖分解为两分子丙酮酸,并且发生氧化(脱氢)和生成少量ATP。
三羧酸循环:丙酮酸彻底分解为CO2和氢(这个氢被传递氢的辅酶携带着),同时生成少量的ATP。
氧化磷酸化:氢(氢离子和电子)被传递给氧以生成水,并且放出大部分的能量,以生成ATP。
(1)糖酵解
糖酵解是葡萄糖氧化的第一阶段。包括一系列反应,都是在细胞质中发生的,不需要氧,每一步反应都有特定的酶催化。
糖酵解的全过程,主要包括以下步骤:
①葡萄糖磷酸化:葡萄糖氧化是放能反应,但葡萄糖是比较稳定的化合物,如果要使它氧化放出能量来,必须先给予活化能来推动这个反应,使葡萄糖从稳定状态变为活跃状态。这个活化能是由ATP提供的。一个ATP的磷酸通过己糖激酶的催化反应而连到葡萄糖分子的6位碳上,使葡萄糖成为葡萄糖–6–磷酸,这一反应是放能反应,一个ATP放出一个高能磷酸键,大约释放出30.5kJ自由能,其中大部分变为热而散失,小部分用于使磷酸与葡萄糖结合,由于葡萄糖中的磷酸键不是高能的,所以写成直线。
②葡萄糖–6–磷酸经异构酶的催化而变为它的异构体果糖–6–磷酸,然后又有一个ATP分解,一个磷酸根连到1位C上,成为果糖一1,6–二磷酸。这一反应是通过磷酸果糖激酶的催化而实现的。
反应至此,消耗了2个ATP分子,经过一系列酶的催化一个葡萄糖分子形成一个果糖一1,6–二磷酸分子。
③醛缩酶催化果糖一1,6–二磷酸裂解,产生2个分子的三碳化合物,它们分别是磷酸二羟丙酮和3–磷酸甘油醛(PGALd),以参加进一步的代谢。
以上从一个分子葡萄糖转化为2分子的PGALd,是糖酵解的第一阶段。这一阶段不但没有产生ATP,反而从细胞贮备中消耗了2个ATP。
④PGALd的氧化和磷酸化,生成1,3–二磷酸甘油酸。2个PGALd氧化,脱下2个已同时放出能(-43.1kJ),H由辅酶NAD+(电子受体)接受而成为还原态的NADH+H+;产生的一部分能则贮存于高能磷酸键中,即PGALd分子从细胞质基质的无机磷酸盐(Pi)中吸收一个磷酸,生成1,3–二磷酸甘油酸(DPGA)。由于这一分子中新形成的键是高能的,所以用曲线表示。
⑤DPGA的高能磷酸键转移至ADP,产生ATP和3–磷酸甘油酸,这一反应称为底物水平的磷酸化,以示与氧化磷酸化的区别。至此,细胞从一个分子的葡萄糖获得了2个ATP,同时有2个NAD+还原为2个NADH+H+。
⑥3–磷酸甘油酸经2–磷酸甘油酸转化为磷酸烯醇式丙酮酸,它的磷酸键吸收了自由能而变成了高能键,在丙酮酸激酶的催化下,发生第二次底物水平的磷酸化,又生成2个ATP和2个丙酮酸。
以上是糖酵解的第二阶段,这一阶段共产生了4个ATP。
糖酵解过程的总账是:一分子的葡萄糖分解为2个分子的丙酮酸,NAD+被还原,产生了2个 NADH+H+。总反应为:
葡萄糖+2ADP+2Pi+2NAD+→2丙酮酸+2ATP+2NADH+2H++2H2O
在葡萄糖到丙酮酸的整个过程中,能位是逐步下降的,但只有上述两个反应的能位下降较大,足以生成ATP。其他反应则只有微小的下降,还不足以生成ATP。
葡萄糖经过精酵解过程只放出了不足l/4的化学能,大部分能量还保存在2个丙酮酸分子和2个NADH中。糖酵解发生在胞质中,而丙酮酸的继续氧化包括三羧酸循环和电子传递两个过程,则是在线粒体中进行的。丙酮酸在进入三羧酸循环之前,先要氧化脱羧,与辅酶A结合成为活化的乙酸辅酶A(简写为乙酸CoA)。这一过程除释放出1分子CO2外(这是细胞呼吸最早释放出来的CO2),同时还发生NAD+的还原。氧化脱羧实际上就是脱氢脱羧过程。丙酮酸的氧化脱羧是在线粒体基质中进行的,所产生的乙酸CoA即进入三羧酸循环(或称柠檬酸循环),三羧酸循环也发生在线粒体基质中。
(2)柠檬酸循环
柠檬酸循环的得名是由于在这个循环中有一个关键的中间代谢物,即柠檬酸。柠檬酸是一种三羧酸,所以这一循环又称为三羧酸循环(简称为TCA循环)。
柠檬酸循环途径中的酶,除琥珀酸脱氢酶定位于线粒体内膜之外,其余均存在于线粒体基质中。柠檬酸循环的全过程分述如下:
柠檬酸循环的第一步是每个二碳的乙酰CoA分子和一个四碳的草酰乙酸分子结合,生成六碳的柠檬酸:
乙酸CoA+草酰乙酸→柠檬酸+CoA
柠檬酸继续氧化,逐步脱去2个羧基,生成四碳化合物,最后又形成四碳的草酰乙酸,再与乙酰CoA结合,开始另一次循环。在这一全过程中,丙酮酸的3个碳在转变为乙酰CoA时脱去一个,在柠檬酸循环中脱去2个。这3个碳原子氧化的结果生成3个分子CO2,这样葡萄糖中的碳就被完全氧化了。
TCA过程中发生的主要事件的顺序为:
① 乙酰CoA与草酰乙酸结合,生成六碳的柠檬酸,释放CoA。
② 柠檬酸分子不能进行脱氢反应,它先失去一个H2O而成为顺乌头酸,再结合一个 H2O转变为异柠檬酸。
③ 异柠檬酸同时进行脱氢和脱数反应,生成五碳的α–酮戊二酸,放出一个CO2,同时一个NAD+还原为NADH+H+。
④ a–酮戊二酸也同时进行脱氢和脱羧反应,并和CoA结合,生成含有一个高能硫键的四碳化合物,即琥珀酸CoA,同时释放出一个CO2,并且将NAD+还原为NADH+H+。故这一反应与丙酮酸的脱氢脱羧反应相同,也是有CoA参与的。
⑤ 琥珀酰CoA脱去CoA和高能硫键而成为琥珀酸,放出的能量则转入高能磷酸键中,即生成三磷酸鸟苷(GTP),GTP再将高能磷酸转入ADP产生ATP。这也是底物水平磷酸化的过程。
⑥ 琥珀酸脱氢生成延胡索酸。催化这一反应的琥珀酸脱氢酶定位于线粒体内膜上,其辅酶是FAD,而不是NAD+,所以在这里是FAD接受氢而生成FADH2。
⑦ 延胡索酸和水化合而成苹果酸。
⑧ 苹果酸氧化脱氢,生成草酰乙酸,亦即草酰乙酸再生,可重新与新的乙酸CoA分子结合,开始新一轮循环。在这一反应过程中,一个NAD+还原为NADH+H+。
柠檬酸进行一次循环,投入的原料是二碳的乙酸酰CoA,并释放2分子CO2,8个氢(8个质子和8个电子),其中4个来自乙酰CoA,另4个来自加入的水分子。这些氢被传递到电子受体上,生成3分子NADH+H+和1分子FADH2。此外,柠檬酸循环中还生成了1分子ATP,这也属于底物水平的磷酸化。由于一个葡萄糖分子产生2个乙酰CoA,所以一个葡萄糖分子在柠檬酸循环中共产生4个CO2分子6个NADH+H+分子个FADH2分子和2个ATP分子。
(3)电子传递系统和氧化磷酸化
葡萄糖经过糖酵解和柠檬酸循环而全部被氧化,氧化所产生的能量一部分储存在ATP中,一部分还保留在NADH和FADH2中。NADH+H+和FADH2中的能量如何释放出来而转移给ATP呢?这是靠包括分子氧在内的电子传递系统或电子传递链来完成的。
电子传递链就是存在于线粒体内膜上的一系列电子传递体,如FMN、CoQ和各种细胞色素等。分子氧是电子传递链中最后的电子受体。糖酵解和柠檬酸循环产生的NADH+H+和FADH2中的电子和H+,沿着电子传递链上各电子传递体顺序地传递,最后到达分子氧。在这一过程中,所释放的能量就通过磷酸化而被储存到ATP中。所以这里的ATP的形成是发生在线粒体内膜上。这里发生的磷酸化作用是和氧化过程的电子传递紧密相关的,所以和底物水平磷酸化不同,称为氧化磷酸化。关于氧化磷酸化的机制,目前公认的是化学渗透学说。该学说是英国人P.Mitchell经过大量实验后于1961年首先提出的,其主要论点是认为呼吸链存在于线粒体内膜之上,当氧化进行时,呼吸链起质子泵作用,质子被泵出线粒体内膜之外侧(膜间隙),造成了膜内外两侧间跨膜的电化学势差,后者被膜上ATP合成酶所利用,使ADP与Pi合成ATP。每2个质子顺着电化学梯度,从膜间隙进入线粒体基质中所放出的能量可合成一个ATP分子。一个NADH+H+分子经过电子传递链后,可积累6个质子,因而共可生成3个ATP分子;而一个 FADH2分子经过电子传递链后,只积累4个质子,因而只可以生成2个ATP分子。一个葡萄糖分子经过有氧呼吸的全过程,总共能生成的ATP分子数统计如下:
糖酵解:①底物水平的磷酸化…………………………………4个ATP(细胞质基质)
②已糖分子活化消耗…………………………………2个ATP(细胞质基质)
③产生2分子NADH+H+,经过电子传递生成……4个或6个ATP(线粒体)
净积累………………………………………………6或 8个ATP
丙酮酸氧化脱羧:产生2分子NADH+H+(线粒体),可生成………………6个ATP
柠檬酸循环:①底物水平磷酸化(线粒体)…………………………………2个ATP
②产生6个NADH+H+(线粒体),可生成…………………18个ATP
③产生2分子FADH2(线粒体),可生成……………………4个ATP
总计生成…………………………………………………36或38个ATP
在糖酵解过程中产生的2分子NADH+H+为什么有时生成4个ATP,有时又生成6个ATP呢?这是因为:糖酵解是在细胞质基质中进行的,在真核生物中,细胞质基质中的NADH+H+不能通过正常的线粒体内膜,线粒体外的NADH+H+可将其所带的H转交给某种能透过线粒体膜的化合物,进入线粒体内以后再氧化。胞质中的NADH+H+是通过下列两种穿梭运送的系统而到达线粒体基质,然后再通过线粒体内膜上的呼吸链进行氧化的。
① 磷酸甘油穿梭系统:胞质中含有以NADH+H+为辅酶的磷酸甘油脱氢酶可以将磷酸二羟丙酮还原为磷酸甘油,后者可以扩散到线粒体基质内。线粒体基质内,则有另一种含有FAD的α–磷酸甘油脱氢酶,它可催化进入的1–磷酸甘油脱氢,形成FADH2。于是胞质内的NADH+H+便间接地形成了线粒体基质内的FADH2,后者通过呼吸链产生ATP,每1分子FADH2可产生2分子ATP。这种穿梭作用主要存在于肌肉、神经组织中。所以葡萄糖在这些组织中彻底氧化所产生的ATP比其他组织中要少2个,即产生36个ATP。
② 苹果酸–草酰乙酸穿梭系统:胞质中含有苹果酸脱氢酶,可催化草酰乙酸还原为苹果酸,后者可以进入线粒体基质。线粒体基质内则有另一种苹果酸脱氢酶,可催化进入的苹果酸脱氢形成草酰乙酸和NADH+H+,于是胞质内的NADH+H+上的H便间接地被转运进入线粒体基质中。草酰乙酸则通过基质和胞质均含有的谷–草转氨酶的作用,从基质返回胞质中。每1分子NADH进入线粒体内膜的呼吸链氧化可产生3分子ATP。在心脏、肝、肾等细胞中,胞质中的NADH+H+属于此种穿梭。所以1分子葡萄糖在这些组织中彻底氧化所产生的ATP分子数为38个。
2.无氧呼吸的过程
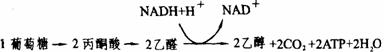
(1)酒精发酵:酵母菌和其他一些生物,甚至一些高等植物,在缺氧条件下,都以酒精发酵的形式进行无氧吸吸。这一过程简单地说就是葡萄糖经糖酵解而成为丙酮酸,丙酮酸脱羧放出CO2而生成乙醛,乙醛被NADH+H+而还原成酒精(见下图)。
(2)乳酸发酵:是某些微生物,如乳酸菌的无氧吸吸过程。高等动物有乳酸发酵过程。人在激烈运动时,氧一时供应不足,葡萄糖酵解产生的部分丙酮酸不能氧化脱羧,因而不能进入三羧酸循环,这时丙酮酸就进入乳酸发酵途径(见下图)。
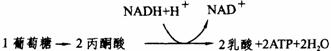
乳酸菌可以使牛奶发酵制成奶酪和酸牛奶。泡菜、酸菜、青贮饲料能够较长时间地保存,也都是利用乳酸发酵积累的乳酸,抑制了其他微生物活动的缘故。
糖酵解是厌氧过程,但不是专性的,在有氧及无氧条件下均能进行。在正常的有氧条件下,通过糖酵解形成的丙酮酸进入线粒体的基质中被完全氧化,形成的NADH+H+通过两种穿梭系统进入线粒体内膜的呼吸链被氧化。在缺氧条件下,NADH+H+不能进入呼吸链。要保持糖酵解继续进行,NADH+H+必需再氧化。这时丙酮酸在细胞质基质中脱羧形成乙醛,乙醛在乙醛脱氢酶作用下,以 NADH+H+作为还原剂形成乙醇,或是丙酮酸在乳酸脱氢酶作用下,直接被NADH+H+还原为乳酸,从而使NADH+H+氧化为NAD+,再接受糖酵解过程脱下的氢。故1分子葡萄糖无论是进行了酒精发酵还是乳酸发酵,均只产生2分子ATP。
(三)光合作用
光合作用大致可分为下列三大步骤:第一步,光能的吸收、传递和转换成电能的过程(通过原初反应完成);第二步电能转变为活跃的化学能过程(通过电子传递和光合磷酸化完成);第三步,活跃的化学能转变为稳定的化学能过程(通过碳同化完成)。第一、二两大步骤基本上属于光反应,第三大步骤属于暗反应(见下表)。

1.原初反应
原初反应包括光能的吸收、传递与转换过程。
根据其功能来区分,叶绿体片层结构上的色素又可区别为两种:一种是作用中心色素,少数特殊状态的叶绿素a分子属于此类,它具有光化学活性,既是光能的“捕捉器”,又是光能的“转换器”(把光能转换为电动势);另一种是聚光色素,没有光化学活性,只有收集和传递光能的作用,能把光能聚集起来,传到作用中心色素,绝大多数色素(包括大部分叶绿素a和全部叶绿素b、β-胡萝卜素、叶黄素、藻红蛋白和藻蓝蛋白)都属于聚光色素。
当波长范围为400-700nm的可见光照到绿色植物上时,聚光系统的色素分子吸收光量子被激发起来。由于叶绿体片层上的色素分子排列得很紧密,光能在色素分子间以诱导共振方式进行传递。能量可以在相同色素分子之间传递,也可以在不同色素分子之间传递。能量传递的效率很高,速度很快,这样就把大量的光能吸收、聚集,并迅速传递到作用中心色素分子,以进行光反应,这个反应部位称为作用中心。光合作用中心至少包括一个光能转换色素分子(P)、一个原初电子受体(A)和一个原初电子供体(D),才能导致电荷分离,将光能转换为电能,并且累积起来。作用中心色素分子一般用其对光线吸收高峰的波长作标志,例如P700代表光能吸收高峰在700的色素分子。作用中心的原初电子受体是指直接接受作用中心色素分子传来电子的物体。作用中心原初电子供体,是指以电子直接供给作用中心色素分子的物体。光合作用的原初反应是连续不断地进行的,因此必须有连续不断的最终电子供体和最终电子受体,构成电子的“源”和“流”。高等植物最终的电子供体是水,最终的电子受体是NADP+。下图表示光合作用的能量吸收、传递与转换的关系。
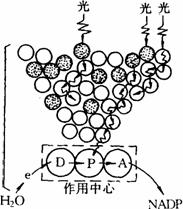
光合作用原初反应的能量吸收、传递与转换图解
粗的波浪箭头是光能的吸收,细的波浪箭头是能量的传递,直线箭头是电子传递。空心圆圈代表聚光性叶绿素分子,有黑点圆圈代表类胡萝卜素等辅助色素。P是作用中心色素分子,D是原初电子供体,A是原初电子受体,e是电子
从图中可以看出,聚光色素分子将光能吸收、传递至作用中心后,使作用中心色素(P)被激发而成为激发态,放出电子给原初电子受体(A),中心色素失去的电子可由原初电子供体(D)来补充,于是中心色素恢复原状,而原初电子供体被氧化。这样不断地氧化还原,就把电子不断地传递给原初电子受体,从而完成了光能转换为电能的过程。
2.电子传递和光合磷酸化
作用中心色素分子被激发后,把电子传递给原初电子受体,转为电能,再通过水的光解和光合磷酸化,经过一系列电子传递体的传递,最后形成ATP和NADPH+H+,从而将电能转化为活跃的化学能,并把化学能贮藏于这两种物质之中。
光合作用的光化学反应是由两个光系统完成的。由于现代研究技术的进展,已经直接从叶绿体中分离出下列两个光系统,即光系统Ⅰ(简称PSⅠ)和光系统Ⅱ(称PSⅡ)。每个光系统均具有特殊的色素复合体及一些物质。光系统Ⅰ的颗粒较小,直径为110埃,位于类囊体膜的外侧;光系统Ⅱ的颗粒较大,直径为175埃,位于类囊体膜的内侧。PSⅠ的光反应是长波光反应,其主要特征是NADP+的还原,其作用中心是P700。当PSI的作用中心色素分子P700吸收光能而被激发后,把电子供给 Fd(铁氧还蛋白),在NADP还原酶的参与下,Fd把NADP+还原成NADPH+H+。PSⅡ的光反应是短波光反应,其主要特征是水的光解和放氧。光系统Ⅱ的作用中心色素分子可能是P680,它吸收光能,把水分解,夺取水中的电子供给光系统Ⅰ。连接着两个光系统的电子传递链,是由一系列互相衔接着的电子传递物质(光合链)组成的。光合链中的电子传递体是质体醌(PQ)、细胞色素b559、Cytf和质体蓝素(PC)等。关于两个光系统的光化学反应和电子传递,如图下图所示。
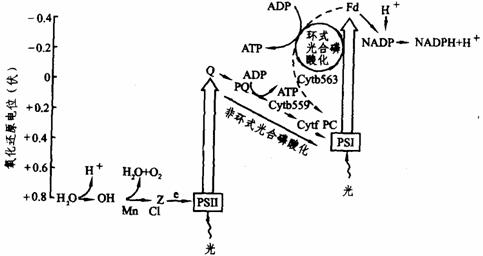
光合作用中的两个光化学反应和电子传递
Z–原初电子供体 Q–未知因素 Fd–含铁氧化还原蛋白(-0.43伏)
光合作用中,磷酸化和电子传递是偶联的,在光反应的电子传递过程中能产生ATP,即叶绿体在光作用下把无机磷和ADP转化成ATP,形成高能磷酸键,此称为光合磷酸化。光合磷酸化又分为非循环式光合磷酸化和循环式光合磷酸化两种类型。
光系统Ⅱ所产生的电子,即水光解释放出的电子,经过一系列的传递,在细胞色素链上引起了ATP的形成。同时把电子传递到PSⅠ上去,进一步提高能位,使H+还原 NADP+成为NADPH+H+。在这个过程中,电子传递不回到原来的起点,是一个开放的通路,故称非循环式光合磷酸化。其反应式为:
2ADP+2Pi+2NADP++2H2O 2ATP+2NADPH+2H++O2
2ATP+2NADPH+2H++O2
光系统Ⅰ产生的电子经过铁氧还蛋白和细胞色素b563等后,只引起ATP的形成,而不放氧,不伴随其他反应。在这个过程中,电子经过一系列传递后降低了位能,最后经过质体蓝素重新回到原来的起点,也就是电子的传递是一个闭合的回路,故称为循环式光合磷酸化。其反应式为:
ADP+PiATP
经过光反应后,由光能转变来的电能暂时贮存在ATP和NADPH中。叶绿体用ATP和NADPH+H+,便可在暗反应中同化二氧化碳,形成碳水化合物。因此有人把ATP和NADPH+H+称为还原力或同化力。还原1分子CO2,需要2个NADPH+H+和3个ATP,这3个ATP中有2个产生于非循环式光合磷酸化,还有1个产生于环式光合磷酸化。
3.碳的同化
从能量转换角度来看,碳同化是将ATP和NADPH+H+中的活跃的化学能,转换为贮存在碳水化合物中的稳定化学能。光合作用中,由CO2到己糖的总反应式可表示如下:
6CO2+18ATP+12NADPH+12H++12H2O→6–磷酸果糖(已糖)+18ADP+12NADP++17H3PO4
高等植物光合同化CO2的生化途径有卡尔文循环、C4途径和景天科酸代谢三种。其中以卡尔文循环最基本、最普遍,同时也只有这种途径具备合成淀粉等产物的能力。其他两种不够普遍,而且只能起固定、转运CO2的作用,单独不能形成淀粉等产物,所固定的CO2在植物体内再次释放出来,参与卡尔文循环。
(1)卡尔文循环:卡尔文循环是所有植物光合作用碳同化的基本途径,它能形成碳水化合物并输送到细胞质中。在这个循环中,由于大多数植物还原CO2的第一个产物是三碳化合物(如磷酸甘油酸),故又称为C3途径。卡尔文循环大致可分为核化、还原和再生三个阶段。
①羧化阶段:1,5–二磷酸核酮糖十CO2→3–磷酸甘油酸
②还原阶段:3–磷酸甘油酸→3–磷酸甘油醛
③再生阶段:3–磷酸甘油醛→6–磷酸果糖→5–磷酸核酮糖→1,5–二磷酸核酮糖(简称RuBP)
在此循环途径中,首先是RuBP在核酮糖二磷酸羧化酶催化下与CO2结合,生成3–磷酸甘油酸;3–磷酸甘油酸经磷酸化和脱氢两步反应,生成3–磷酸甘油醛;3–磷酸甘油醛分别经两条途径又重新回到RuBP,继续进行CO2的固定、还原等一系列反应,使循环反复进行。
卡尔文循环的产物不是葡萄糖,而是三碳的丙糖,即3–磷酸甘油醛(简写为PGALd),再由2个PGALd化合而成葡萄糖。这一循环的总账是:循环3次,固定3个CO2分子,生成6个PGALd,其中1个PGALd用来合成葡萄糖或其他糖类,这1个PGALd才是本循环的净收入,其余5个PGALd则用来产生3个分子的RuBP以保证再循环。所以每产生1分子葡萄糖需要2个分子的PGALd,即需要完成6次循环。从能量的变化来计算:生产一个可用于细胞代谢和合成的PGALd,需要9个ATP分子和6个NADPH分子参与。即:
3RuBP+3CO2 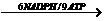 PGALd+3RuBP
PGALd+3RuBP
PGALd在叶绿体中不能积累,需通过一系列转化形成淀粉,作为光合作用的产物,暂时贮存于叶绿体中,或输出叶绿体,在细胞质中转变为蔗糖。一般以淀粉和蔗糖作为光合作用的产物。
(2)C4途径:有些起源于热带的植物,如甘蔗、玉米、高梁等,除了和其他植物一样具有C3途径外,还有一条固定CO2的途径和C3途径联系在一起。这个途径的CO2受体是磷酸烯酸式丙酮酸,在叶肉细胞质中,在磷酸烯酸式丙酮酸(简写为PEP)羧化酶的催化下,固定CO2而生成草酰乙酸。由于还原CO2的第一个产物草酰乙酸是四碳化合物,所以这个途径叫C4途径。具有C4途径的这类植物叫C4植物。
C4植物叶片的结构很独特,含有2种不同类型的光合细胞,各具不同的叶绿体。围绕着维管束鞘细胞周围的排列整齐致密的叶肉细胞中的叶绿体,具有发达的基粒构造,而维管束鞘细胞的叶绿体中却只有很少的基粒,而有很多大的卵形淀粉粒。
在C3植物中,CO2是在叶肉细胞中通过卡尔文循环而被固定还原的。在C4植物的叶肉细胞中,CO2的接受体不是C3途径的RuBP,而是PEP。催化这一反应的酶是PEP羧化酶。CO2被固定后,不是生成三碳的磷酸甘油酸(简写成PGA),而是生成四碳的双羧酸,即草酸乙酸,草酸乙酸再被NADPH还原而成苹果酸。苹果酸离开叶肉细胞,进入维管束鞘细胞中,脱羧放出CO2,而成为丙酮酸。丙酮酸再回到叶肉细胞中,被转变为PEP,继续固定CO2。而苹果酸脱羧产生的CO2,在维管束鞘细胞中仍为RuBP所固定,而进入卡尔文循环。C4植物既有C4途径又有C3途径,这2个途径的关系如下图所示。
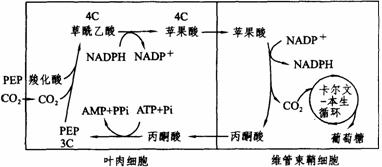
C4植物中的C4途径与C3途径的关系
在C4植物中,CO2在叶肉细胞中先按照C4途径被固定,然后在维管束鞘细胞中仍旧是通过卡尔文循环而被还原。由于在C4植物的C4途径中,PEP羧化酶对CO2的亲和力极强,甚至当CO2浓度降低时,也能固定CO2。所以C4途径是在 CO2浓度低时获取CO2的一种途径。生活在高强光和热带地区的多种植物,气孔经常是关闭的,这样可防止水分的过度散失,但同时也导致体内CO2浓度的降低。C4途径的存在,使CO2不致成为光合作用的限制因子,从而提高了光合效率。这通常是C4植物的生产效率明显高于C3植物的重要原因之一。C3植物生产效率较低的另一个原因是它们具有较强过程的光呼吸。
4.光呼吸
光呼吸是指绿色植物只在光照条件下才能吸收氧气,放出CO2的过程。光呼吸和一般生活细胞的呼吸作用(通过线粒体释放CO2的呼吸作用)显著不同,它是在光刺激下绿色细胞释放CO2的现象。光呼吸的高低。是指植物在光合作用下释放CO2的多少,这样释放的CO2,实际上是植物在光合作用过程中同化的CO2,它往往将光合作用已固定的20%- 40%的碳变成CO2再释放出来。显然这是一个消耗过程,对积累光合产物很不利。
光呼吸的底物是乙醇酸。乙醇酸来自叶绿体,叶绿体中的RuBP羧化酶既是羧化酶,催化CO2与RuBP结合,又是加氧酶,催化O2与RuBP结合。在CO2分压低、氧分压高时,这个酶催化O2与RuBP结合而生成三碳的3–PGA和二碳的2–磷酸乙醇酸。2–磷酸乙醇酸水解而成乙醇酸和无机磷酸。乙醇酸进入过氧化物体,在这里被氧化,其产物进入线粒体,在这里释放出CO2,这就是光呼吸的全过程。
(二)矿质元素的吸收和运输
1.植物细胞对矿质元素的吸收
植物细胞吸收矿质元素的方式有三种:被动吸收、主动吸收和胞饮作用。其中被动吸收和主动吸收是植物细胞吸收矿质元素的主要方式。
(1)被动吸收
被动吸收:是指由于扩散作用或其他物理过程而进行的吸收,是不需要消耗代谢能量的,故又称非代谢性吸收。
①简单扩散:扩散作用是指气体分子、溶剂分子、溶质分子从浓度高的部位向低浓度的部位均匀分布的趋向。当外界溶液的浓度大干细胞内部溶液浓度时,外界溶液中的溶质便扩散进入细胞内,当细胞内外的浓度差大时,细胞大量吸收物质,但随着浓度差变小,吸收也随之减少,直至细胞内外浓度达到平衡为止。所以浓度差是决定细胞靠扩散作用吸收物质的主要因素。
②杜南平衡:杜南平衡是一种特殊的积累离子的现象。杜南平衡的结果是膜两侧某离子的浓度不相等,但也达到了平衡。植物细胞的质膜是一种半透膜。细胞内含有许多带电荷的不能扩散到细胞外的大分子化合物(如蛋白质,R-),成为不扩散离子,它们可以与阳离子形成盐类(如蛋白质的钾盐,KR),设其浓度为Ci,把这样的细胞放在浓度为C。的KCl溶液中,由于细胞内没有Cl-,所以Cl-沿着浓度梯度由外界溶液扩散入细胞内,同时K+也进入细胞内,以保持电中性。由于R-不能向细胞外扩散,使得细胞内的K+被保留在细胞内。经过一段时间后,细胞内外离子扩散速度相等,达到平衡状态,此时,细胞内可扩散负离子和正离子浓度的乘积等于细胞外正负离子浓度乘积。即:
[K+]内×[Cl-]内=[K+]外×[Cl-]外
由于细胞内有部分可扩散正离子被不扩散的负离子吸引,所以扩散平衡时,细胞内K+浓度大于细胞外K+的浓度,呈现离子积累现象,此时细胞外Cl-的浓度大于细胞内Cl-浓度。这种离子积累不需要消耗能量。
(2)主动吸收
在生物膜上,有一类专门运送物质的蛋白质大分子(称运输酶、或透过酶),能有选择性的把膜外的物质透过膜送到膜内,也可把膜内物质运送到膜外。这种物质转运有两个特点:①需要消耗呼吸作用所提供的能量,且被转运的离子或分子数量与所消耗的能之间有一定的量的关系;②转运的速度超过扩散的速度;当转运达到最终的稳衡状态时,膜两侧物质的浓度不相等。这种利用呼吸释放的能量做功而逆着浓度梯度快速吸收离子的过程称为细胞的主动吸收。凡是影响呼吸作用的因素,都会影响细胞的主动吸收。
2.植物吸收矿质元素的特点
(1)离子的选择吸收
植物从环境中吸收离子时是具有选择性的,即吸收离子的数量不与环境溶液中离子浓度成正比。例如,池塘水中K+的浓度较低,但生长在池中的丽藻液泡中积累的K+浓度很高。海水中Na+浓度很高,但生长在海水中的法囊藻液泡中Na+浓度比较低。这种选择性吸收不仅表现在对不同的盐分吸收量不同,而且对同一盐的阳离子和阴离子吸收量也不相同。例如,供给(NH4)2SO4时,根对NH4+吸收多于SO42-,所以溶液中留有许多,使溶液的氢离子浓度增大,这种盐称为生理酸性盐。大多铵盐属于这类盐。相反,NaNO3和Ca(NO3)2属生理碱性盐。此外,还有一类化合物的阴离子和阳离子几乎以同等速度被极吸收,对土壤溶液的酸碱性不产生影响,这类盐称为生理中性盐,如NH4NO3。
(2)单盐毒害和离子对抗
如果将植物培养在只含一种金属离子的溶液中,即使这种离子是植物生长发育所必需的,如K+,而且在培养液中的浓度很低,植物也不能正常生活,不久即受害而死。这种由于溶液中只含有一种金属离子而对植物起毒害作用的现象称为单盐毒害。如果 在能引起毒害作用的溶液中加入另一种矿质离子,其对植物的毒害作用即能减弱或消失,这种离子间能够相互消除毒害的现象称为离子对抗。如在KCl溶液中加入少量的 Ca2+,则K+对植物就不产生毒害作用,即Ca2+能对抗K+。植物只有在含有适当比例的多种盐的溶液中,才能很好的生长,因为这时各种离子的毒害作用已基本上被消除。也就是说,这种溶液既能保证植株有良好的充足的矿质营养,又对植物无毒害效应,这种溶液就称为平衡溶液。相对来说,土壤溶液就是平衡溶液。
值得注意的是:并不是任何一种其他离子的存在都能对抗由于某一种离子而引起的毒害作用。如K+、Na+之间就不存在对抗作用,因为二者对原生质的影响是一致的。而Ca2+与它们的作用正相反,故Ca2+与K+、Na+可以产生拮抗。同样Ba2+不能对抗Ca2+和Mg2+,原理也在于此。
3.矿质元素在植物体内的运输
根部吸收的矿质元素,有一部分留在根内,大部分运输到植物体的其他部分。根部吸收的无机氮大部分在根内转变为有机氮化合物。氮的主要运输形式是氨基酸(天冬氨酸)和酪氨(天冬酰氨和谷氨酰氨),还有少量以NO3-形式向上运输。磷主要以H2PO4-或HPO42-形式运输,也有少量磷在根部转变为有机磷向上运输。硫的运输形式主要是SO42-,也有少量硫以蛋氨酸或谷胱甘肽之类的有机硫形式运输。金属离子都以离子状态运输。
根部吸收的矿质元素进入导管后,沿木质部随着蒸腾流向上运输。叶片吸收的矿质元素在茎部主要是经韧皮部向上向下运输,也可从韧皮部横向运输到木质部,再向上运输。
有些矿质元素进入植物体后形成难溶解的稳定化合物,而被固定下来,不能移动,只能利用一次。最典型的元素是钙,其次是铁、锰等。这类元素在植物体内的分布特点是器官越老含量越高。有些元素在植物体内可多次利用,即该类元素形成化合物后,又可被分解,运输到其他需要的器官去。被再利用的元素中以氮、磷最为典型。
(一)水分的吸收、运输
1.自由能和水势
当把一小块高锰酸钾结晶投入到一盛有纯水的烧杯中时,高锰酸钾分子会迅速地由结晶处向烧杯中的其他地方迁移。这种迁移之所以能够发生,完全是由于结晶与烧杯中的其他地方存在着化学势差的结果。化学势就是在恒温恒压条件下,一摩尔的物质分子所具有的自由能,自由能则是在恒温恒压条件下能够用于做功的能量。所以化学势就是指物质分子能够用于做功的能量的度量。其大小与物质的浓度或纯度呈正相关关系,并且能够指示物质分子发生反应或产生运动的方向和限度。在上述系统中,高锰酸钾分子迁移消耗的就是高锰酸钾分子的化学势或者说就是高锰酸钾分子的自由能。正因为如此,高锰酸钾分子也只能由化学势较高的结晶向化学势较低的其他地方迁移,直到烧杯各处高锰酸钾的化学势都相等为止。这种物质分子顺着化学势梯度或浓度迁移的现象就叫扩散。化学势用μ来表示,单位是耳格/摩尔或达因·厘米 / 摩尔。
在上述系统中高锰酸钾分子扩散的同时,水分子也在扩散,消耗的是水的化学势,是水中能够用于做功的能量度量。其大小当然能够指示水分子发生反应或产生运动的方向和限度,包括植物体内的水分运动。但是,任何物质分子的化学势的绝对值并不容易测定,水的化学势亦如此。我们通常所说的水的化学势实际上是一个差值,是系统中水的化学势与0℃、1.013×105 Pa下纯水的化学势之差。尽管纯水的化学势的绝对值也不易测定,但人们可以规定一个值来作为纯水的化学势,其他溶液的水的化学势就通过与纯水的化学势的值进行比较而得到。
但是,在植物生理学上,一般并不以水的化学势差的大小来指示水分运动的方向和限度,而是以水势的大小来指示的。水势就是每偏摩尔体积水的化学势差,就是水的化学势差被水的偏摩尔体积来除所得的商。即:
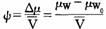
式中Ψ表示的是水势;μw表示的是系统中水的化学势;μw0表示的是纯水的化学势;Δμ表示的是化学势差; 表示的是系统中水的偏摩尔体积。现举一例来说明。
表示的是系统中水的偏摩尔体积。现举一例来说明。
在20℃、1.013×105Pa下,一摩尔纯水的体积是18.09mL,一摩尔纯乙醇的体积是58.35mL,但二者相混合后的体积并不等于18.09+58.35=76.44mL,而是74.40mL。就是说在这个系统中,一摩尔的水所体现出来的体积已不再是18.09mL(而是17.0mL),一摩尔的乙醇所体现出来的体积也不再是58.35mL(而是57.40mL)。这17.0mL和57.40mL分别是水和乙醇在此混合物中此条件下的偏摩尔体积。
水势的大小决定于化学势差的大小,纯水的化学势最大,并规定在0℃、1.013×105Pa下为0,所以纯水的水势也最大,在0℃、1.013×105Pa下也为0。其他的任何溶液(在开放系统中)都由于溶质的存在,降低了水的自由能而使水的化学势都小于纯水,全为负值。水势当然也比纯水小,也全为负值。
水势的大小能够指示水分发生反应或产生运动的方向和限度,并且与化学势所指示的完全相同,无论在植物体外还是在植物体内,水分总是顺着水势梯度由高水势流向低水势区。水势的单位是压力单位,达因 / 厘米2,这可由ψ=Δμ/ 推出,通常以帕斯卡(pa)来表示。
2.植物细胞的水势与渗透吸水
成熟的植物细胞外为纤维素和果胶质组成的细胞壁,中央有一个大的液泡,细胞壁和液泡之间则是细胞的原生质体。从物质透过角度讲,细胞壁是一个完全的透性膜,水分和溶质都可以自由地透过。而原生质膜和液泡膜则是分别透性膜。而且原生质膜和液泡膜之间的中质也并非是任何物质都容易透过的结构。这样我们就有充分的理由将细胞的整个原生质体(原生质膜、液泡膜和中质)看做是一个分别透性膜。液泡中是具有一定渗透势的溶液,那么植物细胞所处的环境溶液的情况就不外乎三种。即环境溶液的水势高于细胞的水势(高水势液),环境溶液的水势低于细胞的水势(低水势液),环境溶液的水势与细胞的水势相等(等水势液)。但不论处在何种情况下,植物细胞与外界溶液之间都能够发生渗透作用,只不过在第三种情况下,由于细胞内外无水势差的存在,外观上没有水分进出细胞的现象发生。所以说一个成熟的植物细胞与外界环境溶液共同构成了一个渗透系统,能够发生渗透作用。植物细胞以渗透吸水为主,吸水的动力来自细胞内外的水势之差,那么,植物细胞的水势又该如何计算呢?
成熟的植物细胞中央有大的液泡,其内充满着具有一定渗透势的溶液,所以渗透势肯定是细胞水势的组成之一,它是由于液泡中溶质的存在而使细胞水势的降低值。因此又称为溶质势,用ψs 表示。由于纯水的渗透势最大,并规定为0,所以任何溶液的渗透势都比纯水要小,全为负值。当细胞处在高水势溶液中时,细胞吸水,体积扩大,由于细胞原生质体和细胞壁的伸缩性不同,前者大于后者,所以细胞的吸水肯定会使细胞的原生质体对细胞壁产生一种向外的推力,即膨压。反过来细胞壁也会对细胞原生质体、对细胞液产生一种压力,这种压力是促使细胞内的水分向外流的力量,这就等于增加了细胞的水势。这个由于压力的存在而使细胞水势的增加值就称为压力势,用ψp表示。其方向与渗透势相反,一般情况下为正值。此外,细胞质为亲水胶体,能束缚一定量的水分,这就等于降低了细胞的水势。这种由于细胞的胶体物质(衬质)的亲水性而引起的水势降低值就称为细胞的衬质势,以ψm表示。所以说,植物细胞的吸水不仅决定于细胞的渗透势ψs,压力势ψp,而且也决定于细胞的衬质势ψm。一个典型的植物细胞的水势应由三部分组成,即ψw=ψs+ψp+ψm。
从作用效果看,ψs和ψm是使水分由细胞外向细胞内流的力量;ψp则是使水分由胞内向外渗的力量;就是说ψs和ψm的符号与ψp的符号相反,ψs和ψm为负,而ψp为正。
理论上细胞的水势ψw应由ψs、ψp和ψm三部分组成,但ψs、ψp和ψm在细胞水势中所占的比例则是随着细胞的发育时期及细胞所处的状态的改变而变化的。就ψm来讲,干燥种子和未形成液泡的细胞中,ψm是一个很大的负值;而在有液泡的细胞中,由于细胞的衬质部分已被水饱和,ψm等于零或接近于零,其绝对值很小(<0.1),相对于绝对值很大的水势来讲,就十分的微不足道了。因此,在计算有液泡细胞水势的时候,ψm通常可以省掉。即有液泡细胞的水势可以用公式ψw=ψs+ψp进行表示和计算。当ψw低于外界溶液时,细胞即可吸水。
在一般情况下,细胞的ψp为正值,但处于强烈蒸腾环境中的细胞的ψp为负值,而不为正值。细胞蒸腾失水,细胞体积缩小,最后可失去膨压而达到萎蔫的程度,但此时一般并不能引起质壁分离,原因是水与细胞壁的附着力很强,这样在原生质体收缩时细胞壁被向里拉,甚至发生褶皱变形。同时,细胞壁产生的反作用力使原生质体和细胞液处于张力的状态。张力相当于负的压力;在计算水势时应取负值。因此,在产生张力时,细胞的水势将变得比ψs更负。
3.根系吸水及水分沿导管或管胞上升的动力
植物体水分的获得主要借助于根系对土壤中水分的吸收。根系的吸水方式有两种,即主动吸水和被动吸水。主动吸水是由于根本身的生理活动而引起的水分吸收,与地上部分的活动无关,吸水动力是根压。被动吸水由蒸腾作用而引起,而与根系的活动无关,吸水的动力是蒸腾拉力。那么,根压和蒸腾拉力是怎样产生的呢?
关于根压产生的机制现在还不很清楚,一般是用渗透理论来解释。为更好地理解这个理论,必须首先了解植物体的结构。
植物体从空间上可分为三个部分,即共质体、质外体和液泡。共质体是指植物体中所有细胞中活的部分,即是指整个植物体的原生质总体。由于各细胞原生质之间有许多胞间连丝相连,所以共质体是一个连续的系统或体系。质外体则是指细胞壁、细胞间隙和木质部导管等原生质体以外的部分。水分和溶质可以在其中自由扩散。和共质体不同,质外体是不连续的,由于内皮层凯氏带的存在,内皮层就将质外体分隔成为两个区域,其一在内皮层外,包括皮层部分的细胞壁、细胞间隙,这部分可以和土壤溶液之间保持水分和溶质的扩散平衡。另一区域在中往内,包括中柱部分的细胞壁、细胞间隙及成熟的导管。内外两部分质外体之间的水分和溶质的交流,都只有通过内皮层细胞原生质体部分来进行。液泡由于有液泡膜和原生质体隔开,所以它既不属于质外体,也不属于共质体,而且它们也不连续成整体。
渗透理论认为:土壤中含有丰富的离子,在质外体的外部(皮层),离子随土壤溶液进入质外体直至内皮层,溶液中的离子可被活细胞主动吸收,即由质外体进入共质体。在共质体中,这些离子可以通过胞间连丝从一个细胞运至另一个细胞,通过内皮层进入中柱的活细胞。之后细胞中的离子又被动地扩散到导管中,即由共质体进入质外体。其结果,内皮层以外的质外体离子浓度降低,水势增高;而内皮层以内的质外体离子浓度增高,水势降低。这样内外质外体之间就形成了一个水势梯度,于是水经过内皮层的渗透作用而进入中往,进入导管,使导管内产生一种静水压力即根压,水分即沿导管上升。
蒸腾拉力是由于蒸腾作用而产生的。由于蒸腾,靠近气孔下腔的叶肉细胞含水量减少。水势降低,向相邻细胞吸取水分,当相邻细胞水势减低时,转向其相邻细胞吸水,如此依次传递直至向导管吸水。这就犹如造成了一种将导管中的水向上拉的力量,这种由于蒸腾作用产生的一系列水势梯度使导管中水分上升的力量就称为蒸腾拉力。主动吸水和被动吸水并存,但二者在根系吸水过程中的比重却很不相同。一般被动吸水占有很大的比重,主动吸水很少。所以蒸腾拉力是根系吸水和水分沿导管或管胞上升的主要动力。这里就产生了一个问题,蒸腾拉力将导管中的水柱向上拉,同时水柱本身的重力又将水柱向下拖,水柱的两端同时受到上拉下拖两种力量的作用,使水柱处于紧张状态,产生张力,水柱就有发生中断的趋势,而导管中水柱的连续性对根系的吸水和水分上升来讲又是必要的。那么,在这种情况下,导管中的水柱如何能保持连续而不发生中断呢?众所周知,水分子与水分子之间的内聚力很大,可达-300×105Pa,同时水分子与导管或管胞内纤维素分子之间还有强的附着力,它们远远大于水柱的张力(-5--30×105Pa)。所以导管或管胞中的水柱可以保持连续,保证在蒸腾作用进行时木质部中的水分能不断向上运输。这种以水分子的内聚力大于张力来解释水分上升的学说称为内聚力学说,也称为蒸腾--内聚力--张力学说,是19世纪末爱尔兰人迪克松提出的。
根据最新国际生物学奥林匹克竞赛(IBO)纲要和全国中学生生物学竞赛大纲(试行)要求,有关植物生理学的内容主要包括:水分、矿质营养的吸收和运输;光合作用;呼吸作用;蒸腾作用;生长和发育,激素;生殖。上述内容中,原中学生物教学大纲中已有的不再重复,只对其他一些在竞赛中经常遇到但又难于理解的内容作些简要的分析或说明。
26.向日葵所具有的特征是
|
|
花序 |
花冠 |
雄蕊数/每小花 |
种子数/每小花 |
|
A |
伞形花序 |
分开的 |
2 |
2 |
|
B |
头状花序 |
联合的 |
5 |
1 |
|
C |
头状花序 |
分开的 |
5 |
多数 |
|
D |
伞形花序 |
联合的 |
5 |
1 |
25.根据下列十字花树的花图式,写出化程式。(十字花科为胚珠多数,子房2心皮1室,上位)十字花科的花程式为: 。

24.判断下列植物属于哪个植物类群,将编号填入适当的括号内。
①松萝 ②桫椤 ③菠萝 ④石松 ⑤石花菜 ⑥石莼 ⑦石蕊 ⑧卷柏 ⑨桧柏 ⑩马尾松 11狗尾草 12蜈蚣草 13裙带菜 14贯众 15地钱 16铁树 17满江红 18浮萍 19泥炭癣 20金鱼藻
(1)藻类植物( )。
(2)地衣植物( )。
(3)苔藓植物( )。
(4)蕨类植物( )。
(5)裸子植物( )。
(6)被子植物( )。
23.关于各种植物的叙述,正确的是
A 水稻种子的子叶发达,而蚕豆种子的胚乳发达
B 玉米的维管束呈环状排列,向日葵茎的维管束散生于基本组织中
C 椰子树茎秆粗大,为双子叶植物,菊花的茎瘦弱,为单子叶植物
D 百合花有3个花瓣,梅花有5个花瓣
22.菊花的花序属于下列哪种花序类型
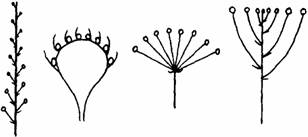
A B C D
湖北省互联网违法和不良信息举报平台 | 网上有害信息举报专区 | 电信诈骗举报专区 | 涉历史虚无主义有害信息举报专区 | 涉企侵权举报专区
违法和不良信息举报电话:027-86699610 举报邮箱:58377363@163.com