题目列表(包括答案和解析)
(一)细胞的生物电现象
生物电现象是指生物细胞膜在安静状态和活动时伴有的电现象。它与细胞兴奋的产生和传导有着密切的关系。现以神经细胞为例来讨论细胞的生物电现象。
1.静息电位及产生原理
(1)静息电位:细胞膜处于安静状态下,存在于膜内外两侧的电位差,称为静息电位。如下图所示,将两个电极置于安静状态下神经纤维表面任何两点时,示波器屏幕上的光点在等电位线作横向扫描,表示细胞膜表面不存在电位差。但如将两个电极中的一个微电极(直径不足1μm)的尖端刺入膜内,此时示波器屏幕上光点迅速从等电位下降到一定水平继续作横向扫描,显示膜内电位比膜外电位低,表示细胞膜的内外两侧存在着跨膜电位差。此电位差即是静息电位。一般将细胞膜外电位看作零,细胞膜内电位用负值表示。
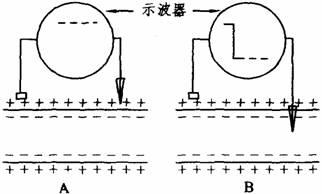
静息电位测量示意图
A.膜表面无电位差 B.膜内外两侧有电位差
同类细胞的静息电位较恒定,如哺乳类动物神经细胞的静息电位为-70--90mV。安静时,细胞膜两侧这种数值比较稳定的内负外正的状态,称为极化。极化与静息电位都是细胞处于静息状态的标志。以静息电位为准,若膜内电位向负值增大的方向变化,称为超极化;若膜内电位向负值减小的方向变化,称为去极化;细胞发生去极化后向原先的极化方向恢复,称为复极化。从生物电来看,细胞的兴奋和抑制都是以极化为基础,细胞去极化时表现为兴奋,超极化时则表现为抑制。
(2)静息电位的产生原理:“离子流学说”认为,生物电产生的前提是细胞膜内外的离子分布和浓度不同,以及在不同生理状态下,细胞膜对各种离子的通透性有差异。据测定,在静息状态下细胞膜内外主要离子分布及膜对离子的通透性见下表。
|
主要离子 |
离子浓度(mmol/L) |
膜内与膜外离子比例 |
膜对离子通透性 |
|
|
膜内 |
膜外 |
|||
|
Na+ K+ Cl- A-(蛋白质) |
14 155 8 60 |
142 5 110 15 |
1︰10 31︰1 1︰14 4︰l |
通透性很小 通透性大 通透性次之 无通透性 |
在静息状态下,由于膜内外K+存在浓度差和膜对K+有较大的通透性,因而一部分K+顺浓度差向膜外扩散,增加了膜外正电荷;虽然膜内带负电的蛋白质(A-)有随K+外流的倾向,但因膜对A-没有通透性,被阻隔在膜的内侧面。随着K+不断外流,膜外的正电荷逐渐增多,于是膜外电位上升,膜内因负电荷增多而电位下降,这样便使紧靠膜的两侧出现一个外正内负的电位差。这种电位差的存在,使K+的继续外流受到膜外正电场的排斥和膜内负电场的吸引,以致限制了K+的外流。随着电位差的增大,K+外流的阻力也随之增大。最后,当促使K+外流的浓度差和阻止K+外流的电位差所构成的两种互相抬抗的力量相等时,K+的净外流量为0,此时跨膜电位就相当于K+的平衡电位。K+的平衡电位与实际测得的静息电位略有差别,通常比测定值略高(即值略小),这是由于在静息状态下,膜对Na+也有较小的通透性,有少量Na+顺浓度差向膜内扩散的缘故。简言之,静息电位主要是K+外流所形成的电一化学平衡电位。
2.动作电位及其产生原理
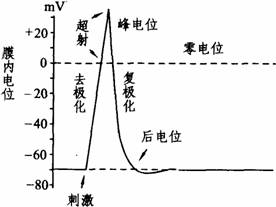 (1)动作电位:细胞膜受到刺激时,在静息电位的基础上发生一次可扩布的电位变化,称为动作电位。动作电位可用上述微电极插入细胞内测量记录下来。在测出静息电位的基础上,给予神经纤维一个有效刺激,此时在示波器屏幕上即显示出一个动作电位(如右图所示)。动作电位包括一个上升相和一个下降相,上升相表示膜的去极化过程,此时膜内原有的负电位迅速消失,并进而变为正电位,即由-70--90mV变为+20-+40mV,出现膜两侧电位倒转(外负内正),整个膜电位变化的幅度可达90-130mV。其超出零电位的部分称为超射。下降相代表膜的复极化过程,是膜内电位从上升相顶端下降到静息电位水平的过程。神经纤维的动作电位,主要部分由于幅度大、时程短(不到2ms),电位波形呈尖峰形,称为峰电位(Spike
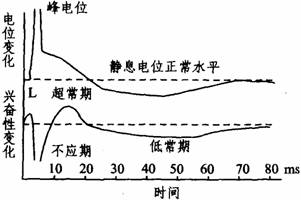
potential)。在峰电位完全恢复到静息电位水平之前,膜两侧还有微小的连续缓慢的电变化,称为后电位。从细胞的生物电角度来看,动作电位与兴奋两者是同义语,而兴奋性是指细胞或组织产生动作电位的能力。动作电位一旦产生,细胞的兴奋性也相应发生一系列改变。从时程上来说,峰电位相当于细胞的绝对不应期;后电位的前段相当于相对不应期和超常期;后电位的后段相当于低常期(如下图所示)。膜电位恢复到静息电位水平,兴奋性也就恢复正常。
(1)动作电位:细胞膜受到刺激时,在静息电位的基础上发生一次可扩布的电位变化,称为动作电位。动作电位可用上述微电极插入细胞内测量记录下来。在测出静息电位的基础上,给予神经纤维一个有效刺激,此时在示波器屏幕上即显示出一个动作电位(如右图所示)。动作电位包括一个上升相和一个下降相,上升相表示膜的去极化过程,此时膜内原有的负电位迅速消失,并进而变为正电位,即由-70--90mV变为+20-+40mV,出现膜两侧电位倒转(外负内正),整个膜电位变化的幅度可达90-130mV。其超出零电位的部分称为超射。下降相代表膜的复极化过程,是膜内电位从上升相顶端下降到静息电位水平的过程。神经纤维的动作电位,主要部分由于幅度大、时程短(不到2ms),电位波形呈尖峰形,称为峰电位(Spike
potential)。在峰电位完全恢复到静息电位水平之前,膜两侧还有微小的连续缓慢的电变化,称为后电位。从细胞的生物电角度来看,动作电位与兴奋两者是同义语,而兴奋性是指细胞或组织产生动作电位的能力。动作电位一旦产生,细胞的兴奋性也相应发生一系列改变。从时程上来说,峰电位相当于细胞的绝对不应期;后电位的前段相当于相对不应期和超常期;后电位的后段相当于低常期(如下图所示)。膜电位恢复到静息电位水平,兴奋性也就恢复正常。
(2)动作电位的引起及产生原理:细胞膜受到刺激后,首先是该部位细胞膜上Na+通道少量开放,膜对Na+的通透性稍有增加,少量Na+由膜外流入膜内,使膜内外电位差减小,称为局部去极化或局部电位,局部电位不能远传。但Na+内流使膜内负电位减小到某一临界数值时,受刺激部位的膜上Na+通道全部开放,使膜对Na+的通透性突然增大,于是膜外Na+顺浓度差和电位差迅速大量内流,从而爆发动作电位。Na+内流是一个正反馈过程(再生性)。使膜对Na+通透性突然增大的临界膜电位数值,称为阈电位。阈电位比静息电位约小10-20mV。任何刺激必须使内负电位降到阈电位水平,才能爆发动作电位。
动作电位上升相是由于膜外Na+大量内流,膜内电位迅速高,使原来的负电位消失并高出膜外电位,在膜的两侧形成一个正外负的电位差。这种电位差的存在,使Na+的继续内流受到内正电荷的排斥,当促使Na+内流的浓度差与阻止Na+内流的位差所构成的两种互相拮抗的力量相等时,Na+的净内流停止。此时膜电位为Na+的平衡电位。简言之,动作电位的上升相是Na+内流所形成的电一化学平衡电位,是膜由K+平衡电位转为Na+平衡电位的过程。
在上升相到达Na+平衡电位时,膜上Na+通道已关闭,Na+的通透性迅速下降。与此同时,膜对K+的通透性大增。于是,K+顺浓度差和顺电位差迅速外流,使膜内外电位又恢复到原来的内负外正的静息水平,形成动作电位的下降相。简言之,动作电位下降相是K+外流所形成,是膜由Na+平衡电位转变为K+平衡电位的过程。
细胞膜在复极化后,跨膜电位虽然恢复,但膜内Na+有所增多,而K+有所减少。这时便激活了细胞膜上的钠一钾泵,通过Na+、K+的主动转运,重新将它们调整到原来静息时的水平,以维持细胞正常的兴奋性。
(3)动作电位的特点:动作电位具有“全或无”现象,刺激达不到阈强度,不能产生动作电位(无),一旦产生,幅度就达到最大值(全)。幅度不随刺激的强度增加而增加。
(4)动作电位的传导特点:动作电位在同一细胞沿膜由近及远地扩布称为动作电位的传导。其传导特点有:①不衰减性传导。动作电位传导时,电位幅度不会因距离增大而减小。②双向性传导。如果刺激神经纤维中段,产生的动作电位可从产生部位沿膜向两端传导。
根据国际生物学奥林匹克竞赛(IBO)纲要和全国中学生生物竞赛大纲(试行)要求,有关人体及动物生理的内容,主要包括消化、吸收、呼吸、循环、排泄、免疫、调节(神经和激素)和生殖。上述内容在中学生物学教学大纲中已有过一些简单的介绍,这里只就竞赛中经常要用到的一些知识作进一步的补充说明。
例1 请用最简便的方法,鉴别核糖、葡萄糖、果糖、蔗糖和淀粉。
分析 鉴别核糖、葡萄糖、果糖、蔗糖和淀粉最简便的方法是显色法。首先在这五种糖中各加入适量碘液,只有淀粉变蓝色,其余四糖不变色。然后在除淀粉外的四糖中分别加适量的盐酸和间苯二酚,核糖呈绿色,葡萄糖呈淡红色,果糖呈红色,而蔗糖不变色。这一下可鉴别出核糖和蔗糖。再在葡萄糖和果糖中分别加入几滴清水,由于葡萄糖具有还原性而使溴水褪色,果糖无还原性,不能使溴水褪色,从而就能达到区分这两种糖的目的。
(四)生物氧化
生物体需要的能量主要是通过代谢物在体内氧化而获得的。物质在生物体内经过氧化最后生成水和CO2并释放能量的过程,称为生物氧化,由于这一过程是在组织细胞内进行的,表现为细胞摄取O2而释放CO2,因此生物氧化又称为组织呼吸或细胞呼吸。虽然糖、脂肪及蛋白质等在体内外氧化分解,都产生水和CO2,并释放出等量的能量,但生物氧化与燃烧的过程有显著不同:生物氧化反应是在温和的条件下进行的酶促反应,所释放的能量,有一部分以化学能的形式储存在ATP分子中。
1.生物氧化的方式和酶类
(1)生物氧化中二氧化碳生成的方式
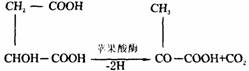
生物体内二氧化碳的生成并不是物质中所含的碳、氧原子的直接化合,而是来源于糖、脂肪等转变来的有机酸的脱羧。根据脱去二氧化碳的羧基在有机酸分子中的位置,可将脱羧反应分为α–脱羧与β–脱羧两种类型。有些脱羧反应不伴有氧化,称为单 纯脱羧;有些则伴有氧化,称为氧化脱羧。例如苹果酸的氧化脱羧:
(2)生物氧化中物质氧化的方式
在化学反应中,失电子、脱氢、加氧都属于氧化;得电子、加氢、脱氧都属于还原。这种变化规律,无论在体内或体外,都是一样的。不同的是,体内氧化都是酶促反应。常见的氧化类型有脱电子、脱氢、加水脱氢和加氧反应。生物体内并不存在游离的电子或氢原子,在上述氧化反应中脱下的电子或氢原子必须为另一物质所接受。这种既能接受又能供出电子或氢原子的物质称为递电子体或递氢体。
(3)与生物氧化有关的酶类
①氧化酶类:能直接利用氧分子作为受氢体,其中氧化酶含铜,反应产物是水,如细胞色素氧化酶。
②需氧脱氢酶类:通常以黄素腺嘌呤二核苷酸(FAD)为辅基,反应产物为过氧化氢。习惯上有时将需氧脱氢酶不严格地称为氧化酶,如黄嘌呤氧化酶。
③不需氧脱氢酶:不能以氧分子而只能以体内某些辅酶为直接受氢体,这些辅酶包括尼克酰胺腺嘌呤二核苷酸(NADP+)、尼克酰胺腺嘌呤二核苷酸磷酸(NAD+)、黄素单核苷酸(FMN)或黄素腺嘌呤二核苷酸(FAD)等。
2.生物氧化中水的生成
生物氧化中水的生成,即代谢物脱下的成对氢原子通过多种酶和辅酶所组成的连锁反应,逐步传递,使之最终与氧结合生成水。由递氢体和递电子体按一定顺序排列构成的此连锁反应与细胞摄取氧的呼吸过程有关,通常称为呼吸链。
(1)呼吸链的组成
呼吸链由许多个组分组成,参加呼吸链的氧化还原酶有尼克酰胺脱氢酶类、黄素脱氢酶类、铁硫蛋白类、细胞色素类、辅酶Q类等。
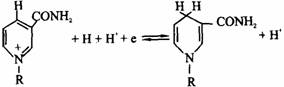
①尼克酰胺脱氢酶类:以NAD+、NADP+为辅酶,它们能够可逆地加氢还原、脱氢氧化,故可作为递氢体而起作用。尼克酰胺只能接受一个氢原子和一个电子,而另一个质子则留在介质中(如下图所示)。
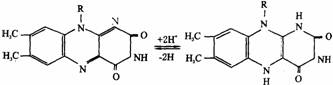
②黄素脱氢酶类:其辅基为FMN、FAD,其分子中都含有异咯嗪。异咯嗪的1和5位的两个氮原子都可以进行脱氢和加氢反应,故它们都可作为递氢体(如下图所示)。
③铁硫蛋白类:其分子中含非卟啉铁和对酸不稳定的硫,其作用是借铁的变价进行电子传递:
Fe3++e===Fe2+
因其分子中含有两个活泼的硫和两个铁原子,故称铁硫中心。
④辅酶Q类:此类酶是一种脂溶性的酯类化合物,因广泛存在于生物界,故又名泛醌。其分子中的苯醌结构能可逆地加氢还原而形成对苯二酚衍生物,故属于传氢体。
⑤细胞色素类:细胞色素是一类以铁卟啉为辅基的蛋白质,在呼吸链中,也依靠铁的化合价的变化而传送电子。目前发现的细胞色素有多种,包括a,a3,b,c,c1等。不同种类的细胞色素的辅基结构及与蛋白质连接的方式是不同的。在典型的线粒体呼吸链中,其顺序是b→c1→c→aa3→O2,其中仅最后一个a3可被氧分子直接氧化,但目前还不能把a和a3分开,故把a和a3合称为细胞色素氧化酶。除aa3外,其余的细胞色素中的铁原子均与其外卟啉环和蛋白质形成六个配位健,惟有aa3的铁原子形成五个配位键,还保留一个配位键,能O2、CO、CN-等结合,其正常功能是与氧结合。
(2)呼吸链中传递体的排列顺序
呼吸链中传递体的排列顺序是根据下列实验数据确定的:
① 根据呼吸键各组分的标准氧化还原电位,按氧还电位递增的顺序依次排列;
② 利用阻断呼吸键的特殊抑制剂,阻断链中某些特定的电子传递环节。若加入某种抑制剂后,则在阻断环节的负电子性侧速电子体(递氢体)因不能再氧化而大多处于还原状态,但在阻断环节的正电子性侧递氢、送电子体不能被还原而大多处于氧化状态。现已基本确定的两条主要的呼吸链中各传递体的排列顺序如下:
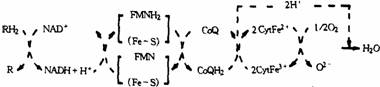
(1)NADH氧化呼吸键(如下图所示)。
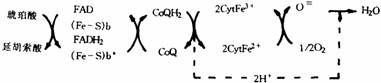
(2)琥珀酸氢化呼吸链(如下图所示)。
(3)胞液中NADH及NADPH的氧化
NADH必须通过线粒体内膜上的呼吸链,其中的氢才能被氧化成水,但是在胞液中形成的NADH(见糖代谢)不能透过正常线粒体内膜,因此线粒体外的NADH尚需通过穿梭系统才能将氢带入线粒体内,而后进行氧化。现已证明,动物体内有下列两种主要的穿梭系统。
①苹果酸穿梭系统(如下图所示)。
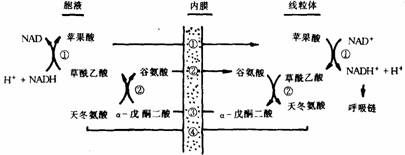
苹果酸穿梭系统
①苹果酸脱氢酶;②谷草转氨酶;①-④线粒体内膜上的不同转位酶
②α–磷酸甘油穿梭系统(如下图所示)。
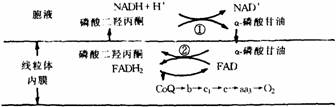
α–磷酸甘油穿梭系统
①胞液中α–磷酸甘油脱氢酶(辅酶为NAD+)
②线粒体内α–磷酸甘油脱氢酶(输基为FAD)
3.ATP与能量的转换和利用
生物体需要利用生物氧化过程中所释放的能量维持生命活动。生物氧化所释放的一部分能量以热能形式散发于周围环境中,约占总能量的60%左右;另一部分则以化学能的形式储存于某些特殊类型的有机磷酸化合物之中。
(1)高能磷酸化合物的生成
在物质氧化过程中大约有40%能量用于ADP的磷酸化合成ATP,该过程有两种方式:一是直接由代谢物分子转移磷酸键至ADP,以合成ATP,即所谓底物水平磷酸化;二是在呼吸键的电子传递过程中偶联的磷酸化,称氧化磷酸化或电子传递水平磷酸化。
(2)氧化磷酸化的偶联部位
根据下述实验数据可以大致确定氧化磷酸化的偶联部位,即ATP的生成部位。
①P/O比值测定及意义:研究氧化磷酸化最常用的方法是测定线粒体的P/O比值。该比值是指每消耗一摩尔氧原子所消耗无机磷的摩尔数,即合成ATP的摩尔数。代谢物脱下的一对氢原子通过NADH呼吸链传递,P/O比值为3;若通过琥珀酸呼吸链传递,P/O比值为2,即分别合成了3或2摩尔ATP。
②根据氧化还原电位之间的电位差计算能量:呼吸链中的三个偶联部位,亦可根据呼吸链传递过程中自由能的变化计算求得,三个ATP分子的形成截获了呼吸链中电子由NADH传递到氧所产生的全部自由能的42%。
(3)氧化磷酸化的抑制作用
①呼吸链阻断剂:能够阻断呼吸链中某一部位电子流的物质称为电子传递阻断剂或呼吸键阻断剂。已知的阻断剂及阻断部。如下图所示。
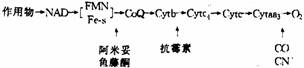
②解偶联剂:解偶联剂对于电子传递没有抑制作用,只抑制由ADP变为ATP的磷酸化作用,即它使产能过程与贮能过程相脱离。2,4–二硝基苯酚是最早发现的一种解偶联剂。
磷酸肌酸是动物体内高能磷酸化合物的贮存形式。在酶的催化下,ATP把高能磷酸键传递给肌酸,以生成磷酸肌酸。当肌肉收缩时,除ATP直接分解供能外,磷酸肌酸又可以转出高能磷酸键,使ADP生成ATP,供组织利用。
(三)酶的特点及功能
酶是由活细胞产生的、具有催化活性和高度专一性的特殊蛋白质。酶被称为生物催化剂。生物体内错综复杂的代谢反应必须具有酶才能按一定规律有条不紊地进行。酶缺陷或酶活性被抑制都会引起疾病。例如,人体缺乏酪氨酸酶会引起白化病。许多中毒性疾病,如有机磷中毒、氰化物中毒、重金属的中毒等,都是由于某些酶的活性被抑制所引起的。
1.酶促反应的特点
酶是生物催化剂,因而它既有与一般催化剂相同的性质,也有与一般催化剂不同的特点。酶和一般催化剂的共同点是:①酶在催化反应加快进行时,在反应前后酶本身没有数量和性质上的改变,因而很少量的酶就可催化大量的物质发生反应。②酶只能催化热力学上允许进行的反应,而不能使本来不能进行的反应发生。③酶只能使反应加快达到平衡,而不能改变达到平衡时反应物和产物的浓度。因此,酶既能加快正反应进行,也能加快逆反应进行。酶促反应究竟朝哪个方向进行,取决于反应物和产物的浓度。酶与一般的催化剂相比又有其特点,最突出的是它的高效性和专一性。
2.酶的化学本质
通过对酶的性质、组成和结构等等方面的研究证实,酶是蛋白质(也有RNA)。蛋白质分为简单蛋白质和结合蛋白质两大类。酶按照化学组成也可分为单纯酶和结合酶两大类。脲酶、胃蛋白酶和核糖核酸酶等一般水解酶都属于简单蛋白质,这些酶只由氨基酸组成,此外不含其他成分。转氨酶、碳酸酐酶、乳酸脱氢酶及其他氧化还原酶等均属于结合蛋白质。这些酶除了蛋白质组分外,还含有对热稳定的非蛋白的小分子物质,前者称酶蛋白,后者称辅因子。酶蛋白与辅因子单独存在时,均无催化活力,只有二者结合成完整的分子时,才具有活力。此完整的酶分子称为全酶(全酶=酶蛋白十辅因子)。有的酶的辅因子是金属离子,有的是小分子有机化合物。通常将这些小分子有机化合物称为辅酶或辅基。辅酶或辅基并没有本质的差别,只不过是它们与蛋白质部分结合的牢固程度不同而已。通常把与酶蛋白结合比较松的,用透析法可除去的小分子有机物称为辅酶;反之为辅基。在酶的催化过程中,辅酶或辅基的作用是作为电子、原子或某些基团的载体参与反应并促进整个催化过程。金属在酶分子中或作为酶活性部位的组成成分,或帮助形成酶活性所必需的构象。一种辅酶常可与多种不同的酶蛋白结合而组成具有不同专一性的全酶。可见决定酶催化专一性的是酶的蛋白质部分。
3.酶的活性中心和必需基团
酶作为蛋白质,其分子比大多数底物要大得多,因此在反应过程中酶与底物的接触只限于酶分子的少数基团或较小的部位。因分子中虽然有许多基团,但并不是所有的基团都与酶的活性有关,其中有些基团若经化学修饰(如氧化、还原、酰化、烷化等)使其改变,则酶的活性丧失,这些基团就称为必需基团。常见的必需基团有Ser的羟基,His的咪唑基,Cys的巯基,Asp、Glu的侧链羧基等。
活性中心(或称活性部位)是指酶分子中直接和底物结合,并和酶催化作用直接有关的部位。对于单纯酶来说,它是由一些氨基酸残基的侧链基团组成的。对于结合酶来说,辅酶或辅基上的某一部分结构往往也是活性部位的组成部分。
酶的活性中心的必需基团可分为两种:一种是与作用物结合的必需基团,称为结合基团,它决定酶的专一性;另一种是促进作用物发生化学变化的基团,称为催化基团,它决定酶的催化能力。但也有些必需基团同时具有这两种作用。另外还有些必需基团位于酶活性中心以外的部位,但仍是维持酶催化作用所必需的,这种称为酶活性中心以外的必需基团。由此可见,酶除了活性中心以外,其他部分并不是可有可无的。活性中心必需基团的作用,一方面使底物与酶依一定构型而结合成为复合物,这样有利于相互影响和作用;另一方面影响底物分子某些键的稳定性,键被打断或形成新的键,从而催化其转变。
某些酶,特别是一些与消化作用有关的酶,在最初合成和分泌时,没有催化活性,这种没有活性的酶的前体称为“酶原”。酶原在一定条件下经适当的物质作用可转变为有活性的酶。酶原转变成酶的过程称为酶原的激活。这个过程实质上是酶活性部位形成或暴露的过程。
4,酶的催化机理
一个反应体系中,任何反应物分子都有进行化学反应的可能,但并非全部反应物分子都进行反应。因为在反应体系中各个反应物分子所含的能量高低不同,只有那些含能量达到或超过一定数值(此能量数值称为此反应的能阈)的分子,才能发生反应,这些分子称为活化分子,使一般分子变为活化分子所需的能量(即分子激活态与基态之间的能量差)称为活化能。在一个反应体系中,活化分子越多,反应就越快,设法增加活化分子的数目就能加快反应的速度。降低活化能,能使本来不够活化水平的分子也成为活化分子,从而增加了活化分子的数目。活化能愈降低,则活化分子的数目就愈多。酶的催化作用就是降低化学反应的活化能,如下图图所示。由于在催化反应中只需较少的能量就可使反应物进入“激活态”,所以同非催化反应相比,活化分子的数量大大增加,从而加快了反应速度。
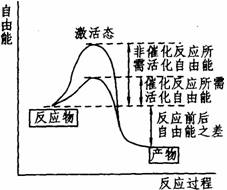
非催化过程与催化过程自由能的变化
目前认为酶降低活化能的原因在于酶参与了反应,即它先与底物结合形成不稳定的中间产物,然后使中间产物再分解,释放出酶及生成反应产物。此过程可用下式表示:

这样,把原来无酶参加的一步反应S===P(能阈较高),变成能阈较低的两步反应(S+E===ES和ES===E+P)。反应的总结果是相同的,但由于反应的过程不同,活化能就大大降低了。这就是目前所公认的中间产物学说。关于酶与底物如何结合形成中间产物,又如何完成催化作用,目前有锁钥学说和诱导契合学说。
5.影响酶作用的因素
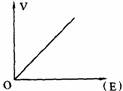 影响酶促反应的因素有酶的浓度、底物浓度、pH值、温度、抑制剂和激活剂等。酶促反应速度指的是反应初速度,此时反应速度与酶的浓度成正比关系,避免反应产物以及其他因素的影响。研究某一因素对酶促反应速度的影响时,在保持其他因素不变的情况下,单独改变研究的因素。
影响酶促反应的因素有酶的浓度、底物浓度、pH值、温度、抑制剂和激活剂等。酶促反应速度指的是反应初速度,此时反应速度与酶的浓度成正比关系,避免反应产物以及其他因素的影响。研究某一因素对酶促反应速度的影响时,在保持其他因素不变的情况下,单独改变研究的因素。
(1)酶的浓度;当底物浓度大大超过酶的浓度,酶的浓度与反应速度呈正比关系(见右图所示)。
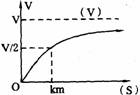 (2)底物浓度:在酶浓度不变的情况下,底物浓度对反应速度影响的作图呈现矩形双曲线(见右图所示)。当底物浓度很低时,反应速度随底物浓度的增加而急骤加快,两者呈正比关系。随着底物浓度的升高,反应速度的增加幅度不断下降。如果继续加大底物浓度,其反应速度不再增加,说明酶已被底物所饱和。所有酶都有饱和现象,只是达到饱和时所需的底物浓度各不相同。
(2)底物浓度:在酶浓度不变的情况下,底物浓度对反应速度影响的作图呈现矩形双曲线(见右图所示)。当底物浓度很低时,反应速度随底物浓度的增加而急骤加快,两者呈正比关系。随着底物浓度的升高,反应速度的增加幅度不断下降。如果继续加大底物浓度,其反应速度不再增加,说明酶已被底物所饱和。所有酶都有饱和现象,只是达到饱和时所需的底物浓度各不相同。
(3)温度:在一定的温度范围内一般化学反应速度均随温度升高而加快,酶促反应也服从这个规律。酶是蛋白质,其本身因温度升高而达到一定高度时会变性,破坏其活性中心的结构,从而减低反应速度或完全失去其催化活性。在某一温度时,酶促反应的速度最大,此时的温度称为酶作用的最适温度。
(4)pH:酶分子中含有许多极性基团,在不同的pH环境中,这些基团的解离状态不同,所带电荷的种类和数量也不尽相同,酶的活性中心往往只处于某一解离状态时最有利于同底物结合。酶催化活性最大时的pH值称为酶作用的最适pH。溶液的pH值高于或低于最适pH时都会使酶的活性降低,远离最适pH值时甚至导致酶的变性失活。
(5)激活剂和抑制剂:激活剂是指能增强酶活性的物质,如Cl-是唾液淀粉酶的激活剂。与激活剂相反,凡能降低酶的活性,甚至使酶完全丧失活性的物质称为酶的抑制剂。抑制剂对酶活性的抑制作用包括不可逆抑制和可逆抑制两类:
①不可逆抑制作用,其抑制剂通常以共价键与酶活性中心上的必需基团相结合,使酶失活。如有机磷化合物能与许多种酶活性中心丝氨酸残基上的羟基结合,使酶失活。
②可逆的抑制作用:包括竞争性抑制与非竞争性抑制两种。在竞争性抑制中,抑制剂常与底物的结构相似,它与底物共同竞争酶的活性中心,从而阻碍底物与酶的结合,如丙二酸对琥珀酸脱氢酶的抑制。非竞争性抑制中的抑制剂可以与酶活性中心外的部位可逆结合,这种结合不影响酶对底物的结合。底物与抑制剂之间无竞争关系,但酶一底物一抑制剂不能进一步释放出产物。对酶促反应速度及其影响因素的研究具有重要的理论和实践意义。
(二)核酸的结构与生物学功能
核酸是生物体内极其重要的生物大分子,是生命的最基本的物质之一。最早是瑞士的化学家米歇尔于1870年从脓细胞的核中分离出来的,由于它们是酸性的,并且最先是从核中分离的,故称为核酸。核酸的发现比蛋白质晚得多。核酸分为脱氧核糖核酸(简称DNA)和核糖核酸(简称RNA)两大类,它们的基本结构单位都是核苷酸(包含脱氧核苷酸)。
1.核酸的基本单位--核苷酸
每一个核苷酸分子由一分子戊糖(核糖或脱氧核糖)、一分子磷酸和一分子含氮碱基组成。碱基分为两类:一类是嘌呤,为双环分子;另一类是嘧啶,为单环分子。嘌呤一般均有A、G2种,嘧啶一般有C、T、U3种。这5种碱基的结构式如下图所示。
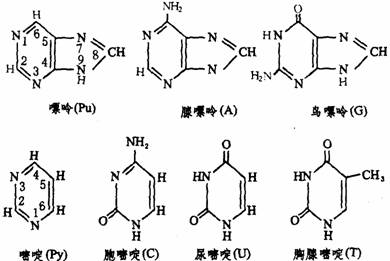
由上述结构式可知:腺嘌呤是嘌呤的6位碳原子上的H被氨基取代。鸟嘌呤是嘌呤的2位碳原子上的H被氨基取代,6位碳原子上的H被酮基取代。3种嘧啶都是在嘧啶2位碳原子上由酮基取代H,在4位碳原子上由氨基或酮基取代H而成,对于T,嘧啶的5位碳原子上由甲基取代了H。凡含有酮基的嘧啶或嘌呤在溶液中可以发生酮式和烯醇式的互变异构现象。结晶状态时,为这种异构体的容量混合物。在生物体内则以酮式占优势,这对于核酸分子中氢键结构的形成非常重要。例如尿嘧啶的互变异构反应式如下图。
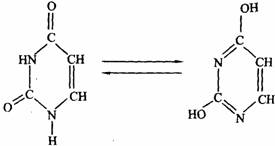
酮式(2,4–二氧嘧啶) 烯酸式(2,4–二羟嘧啶)
在一些核酸中还存在少量其他修饰碱基。由于含量很少,故又称微量碱基或稀有碱基。核酸中修饰碱基多是4种主要碱基的衍生物。tRNA中的修饰碱基种类较多,如次黄嘌呤、二氢尿嘧啶、5–甲基尿嘧啶、4–硫尿嘧啶等,tRNA中修饰碱基含量不一,某些tRNA中的修饰碱基可达碱基总量的10%或更多。
核苷是核糖或脱氧核糖与嘌呤或嘧啶生成的糖苷。戊糖的第1碳原子(C1)通常与嘌呤的第9氮原子或嘧啶的第1氮原子相连。在tRNA中存在少量5–核糖尿嘧啶,这是一种碳苷,其C1是与尿嘧啶的第5位碳原子相连,因为这种戊糖与碱基的连接方式特殊(为C-C连接),故称为假尿苷如下图。
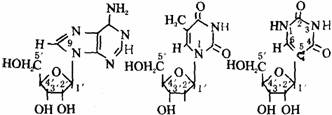
腺苷(A) 脱氧胸苷(dT) 假尿苷(ψ)
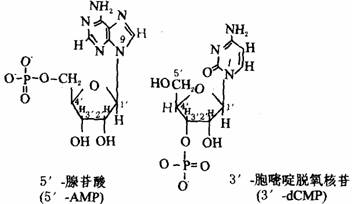
核苷酸是由核苷中糖的某一羟基与磷酸脱水缩合而成的磷酸酯。核苷酸的核糖有3个自由的羟基,可与磷酸酯化分别生成2’–、3’–和5’–核苷酸。脱氧核苷酸的脱氧核糖只有2个自由羟基,只能生成3’–和5’–脱氧核苷酸。生物体内游离存在的核苷酸都是5’–核苷酸。以RNA的腺苷酸为例:当磷酸与核糖5位碳原子上羟基缩合时为5’–腺苷酸,用5’–AMP表示;当磷酸基连接在核糖3位或2位碳原子上时,分别为3’–AMP和2’–AMP。5’–腺苷酸和3’–脱氧胞苷酸的结构式如下图所示。
核苷酸结构也可以用下面简式(如下图)表示。B表示嘌呤或嘧啶碱基,直线表示戊糖,P表示磷酸基。
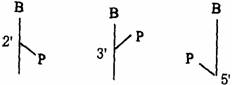
2’–核苷酸 3’–核苷酸 5’–核苷酸
3’–或5’–核苷酸简式也可分别用Np和pN表示(N代表核苷)。即当P在N右侧时为3’–核苷核,P在N左侧的为5’–核苷酸,如3’–核苷酸和5’–核苷酸可分别用Ap和pA表示。
在生物体内,核苷酸除了作为核酸的基本组成单位外,还有一些核苷酸类物质自由存在于细胞内,具有各种重要的生理功能。
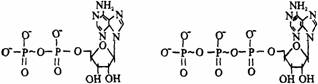
(1)含高能磷酸基的ATP类化合物:5’–腺苷酸进一步磷酸化,可以形成腺苷二磷酸和腺苷三磷酸,分别为ADP和ATP表示。ADP是在AMP接上一分子磷酸而成,ATP是由AMP接上一分子焦磷酸(PPi)而成,它们的结构式如下图所示。
腺苷二磷酸(ADP) 腺苷三磷酸(ATP)
这类化合物中磷酸之间是以酸酐形式结合成键,磷酸酐键具有很高的水解自由能,习惯上称为高能键,通常用“-”表示。ATP分子中有2个磷酸酐键,ADP中只含1个磷酸酐键。
在生活细胞中,ATP和ADP通常以Mg2+或Mn2+盐的复合物形式存在。特别是ATP分子上的焦磷酸基对二价阳离子有高亲和力;加上细胞内常常有相当高浓度的Mg2+,使ATP对Mg2+的亲和力远大于ADP。在体内,凡是有ATP参与的酶反应中,大多数的ATP是以Mg2+-ATP复合物的活性形式起作用的。当ATP被水解时,有两种结果:一是水解形成ADP和无机磷酸;另一种是水解生成AMP和焦磷酸。ATP是大多数生物细胞中能量的直接供体,ATP-ADP循环是生物体系中能量交换的基本方式。
在生物细胞内除了ATP和ADP外,还有其他的5’–核苷二磷酸和三磷酸,如GDP、CDP、UDP和GTP、CTP、UTP;5’–脱氧核苷二磷酸和三磷酸,如dADP、dGDP、 dTDP、dCDP和dATP、dCTP、dGTP、dTTP,它们都是通过ATP的磷酸基转移转化来的,因此ATP是各种高能磷酸基的主要来源。除ATP外,由其他有机碱构成的核苷酸也有重要的生物学功能,如鸟苷三磷酸(GTP)是蛋白质合成过程中所需要的,鸟苷三磷酸(UTP)参与糖原的合成,胞苷三磷酸(CTP)是脂肪和磷脂的合成所必需的。还有4种脱氧核糖核苷的三磷酸酯。即dATP、dCTP、dGTP、dTTP则是DNA合成所必需的原材料。
(2)环状核苷酸;核苷酸可在环化酶的催化下生成环式的一磷酸核苷。其中以3’,5’–环状腺苷酸(以cAMP)研究最多,它是由腺苷酸上磷酸与核糖3’,5’碳原子酯化而形成的,它的结构式如下图所示。
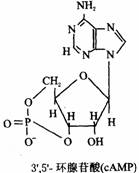
正常细胞中cAMP的浓度很低。在细胞膜上的腺苷酸环化酶和Mg2+存在下,可催化细胞中ATP分子脱去一个焦磷酸而环化成cAMP,使cAMP的浓度升高,但cAMP又可被细胞内特异性的磷酸二酯酶水解成5’–AMP,故cAMP的浓度受这两种酶活力的控制,使其维持一定的浓度。该过程可简单表示如下:
ATP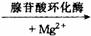 cAMP+焦磷酸
cAMP+焦磷酸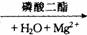 5’–AMP
5’–AMP
现认为cAMP是生物体内的基本调节物质。它传递细胞外的信号,起着某些激素的“第二信使”作用。不少激素的作用是通过cAMP进行的,当激素与膜上受体结合后,活化了腺苷酸环化酶,使细胞内的cAMP含量增加。再通过cAMP去激活特异性的蛋白激酶,由激酶再进一步起作用。近年来发现3’、5’–环鸟苷酸(cGMP)也有调节作用,但其作用与cAMP正好相拮抗。它们共同调节着细胞的生长和发育等过程。此外,在大肠杆菌中cAMP也参与DNA转录的调控作用。
2.核酸的化学结构(或一级结构)
核酸分子是由核苷酸单体通过3’,5’–磷酸二酯键聚合而成的多核苷酸长链。核苷酸单体之间是通过脱水缩合而成为聚合物的,这点与蛋白质的肽链形成很相似。在脱水缩合过程中,一个核苷酸中的磷酸给出一个氢原子;另一个相邻核苷酸中的戊糖给出一个羟基,产生一分子水,每个单体便以磷酸二酯键的形式连接起来。由许多个核苷酸缩合而形成多核苷酸链。如果用脾磷酸二酯酶来水解多核苷酸链,得到的是3’–核苷酸,而用蛇毒磷酸二酯酶来水解得到的却是5’–核苷酸。这证明多核苷酸链是有方向的,一端叫3’–未端,一端叫5’–末端。所谓3’–末端是指多核苷酸链的戊糖上具有3’–磷酸基(或羟基)的末端,而具有5’–磷酸基(或羟基)的末端则称为5’末 –端。多核苷酸链两端的核苷酸为末端核苷酸,末端磷酸基与核苷相连的键称为磷酸单酯键。书写多核苷酸链时,通常将5’端写在左边,3’端写在右边。但在书写一条互补的双链DNA时,由于二条链是反向平行的,因此每条链的末端必须注明5’或3’。通常寡核苷酸链可用右面的简式表示(如右图所示)。
–端。多核苷酸链两端的核苷酸为末端核苷酸,末端磷酸基与核苷相连的键称为磷酸单酯键。书写多核苷酸链时,通常将5’端写在左边,3’端写在右边。但在书写一条互补的双链DNA时,由于二条链是反向平行的,因此每条链的末端必须注明5’或3’。通常寡核苷酸链可用右面的简式表示(如右图所示)。
述简式还可简化为pApCpGpUOH,若进一步简化,还可将核苷酸链中的p省略,或在核苷酸之前加小点,则变为pACGUOH或pA·C·G·UOH。
3.核酸的性质
(1)一般性质
核酸和核苷酸既有磷酸基,又有碱性基团,为两性电解质,因磷酸的酸性强,通常表现为酸性。核酸可被酸、碱或酶水解成为各种组分,其水解程度因水解条件而异。RNA在室温条件下被稀碱水解成核苷酸而DNA对碱较稳定,常利用该性质测定RNA的碱基组成或除去溶液中的RNA杂质。DNA为白色纤维状固体,RNA为白色粉末;都微溶于水,不溶于一般有机溶剂。常用乙醇从溶液中沉淀核酸。
(2)核酸的紫外吸收性质
核酸中的嘌呤碱和嘧啶碱均具有共轭双键,使碱基、核苷、核苷酸和核酸在240-290nm的紫外波段有一个强烈的吸收峰,最大吸收值在260nm附近。不同的核苷酸有不同的吸收特性。由于蛋白质在这一光区仅有很弱的吸收,蛋白质的最大吸收值在280nm处,利用这一特性可以鉴别核酸纯度及其制剂中的蛋白质杂质。
(3)核酸的变性和复性
①核酸的变性:是指核酸双螺旋区的氢键断裂,碱基有规律的堆积被破坏,双螺旋松散,发生从螺旋到单键线团的转变,并分离成两条缠绕的无定形的多核苷酸单键的过程。变性主要是由二级结构的改变引起的,因不涉及共价键的断裂,故一级结构并不发生破坏。多核苷酸骨架上共价键(3’,5’-磷酸二酯健)的断裂称为核酸的降解,降解引起核酸分子量降低。引起核酸变性的因素很多,如加热引起热变性,pH值过低(如pH<4=的酸变性和pH值过高(pH>11.5)的碱变性,纯水条件下引起的变性以及各种变性试剂,如甲醇、乙醇、尿素等都能使核酸变性。此外,DNA的变性还与其分子本身的稳定性有关,由于C-C中有三对氢健而A-T对只有两对氢键,故C+G百分含量高的DNA分子就较稳定,当DNA分子中A+T百分含量高时就容易变性。环状 DNA分子比线形DNA要稳定,因此线状DNA较环状DNA容易变性。
核酸变性后,一系列物理和化学性质也随之发生改变,如260nm区紫外吸收值升高,粘度下降,浮力密度升高,同时改变二级结构,有的可以失去部分或全部生物活性。DNA的加热变性一般在较窄的温度范围内发生,很像固体结晶物质在其熔点突然熔化的情况,因此通常把热变性温度称为“熔点”或解键温度,用Tm表示。对DNA而言,通常把DNA的双螺旋结构失去一半时的温度(或变性量达最大值的一半时的温度)称为该DNA的熔点或解链温度。在此温度可由紫外吸收(或其他特性)最大变化的半数值得到。DNA的Tm值一般在70℃-85℃。RNA变性时发生与DNA变性时类似的变化,但其变化程度不及DNA大,因为RNA分子中只有部分螺旋区。
②核酸的复性:变性DNA在适当条件下,又可使两条彼此分开的链重新缔合成为双螺旋结构,这个过程称为复性。DNA复性后,许多物理、化学性质又得到恢复,生物活性也可以得到部分恢复。DNA的片段越大,复性越慢;DNA的浓度越高,复性越快。
DNA或RNA变性或降解时,其紫外吸收值增加,这种现象叫做增色效应,与增色效应相反的现象称为减色效应,变性核酸复性时则发生减色效应。它们是由堆积碱基的电子间相互作用的变化引起的。
(一)蛋白质的结构和生物学功能
蛋白质是构成细胞和生物体的基本物质,占细胞干重的一半,生物膜中蛋白质的含量占60%-70%,蛋白质在原生质的有机成分中占80%。所有蛋白质的元素组成都很近似,都含有C、H、O、N四种元素,其中平均含氮量约占16%,这是蛋白质在元素组成上的一个特点。蛋白质是一类极为复杂的含氮高分子化合物,其基本组成单位是氨基酸。
1.蛋白质的基本组成单位--氨基酸
组成蛋白质的氨基酸有20种,其中19种结构可用通式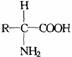 表示。另一种为脯氨酸,它也有类似结构,但侧链与氮原子相接形成亚氨基酸。除甘氨酸外,蛋白质中的氨基酸都具有不对称碳原子,都有L-型与D一型之分,为区别两种构型,通过与甘油醛的构型相比较,人为地规定一种为L型,另种为D一型。当书写时-NH2写在左边为L型,-NH2在右为D一型。已知天然蛋白质中的氨基酸都属L型。
表示。另一种为脯氨酸,它也有类似结构,但侧链与氮原子相接形成亚氨基酸。除甘氨酸外,蛋白质中的氨基酸都具有不对称碳原子,都有L-型与D一型之分,为区别两种构型,通过与甘油醛的构型相比较,人为地规定一种为L型,另种为D一型。当书写时-NH2写在左边为L型,-NH2在右为D一型。已知天然蛋白质中的氨基酸都属L型。
20种基本氨基酸中,有许多是能在生物体内从其他化合物合成的。但其中有8种氨基酸是不能在人体内合成的,叫必需氨酸。它们是:苏氨酸(Thr)、亮氨酸(Leu)、异亮氨酸(Ile)、甲硫酸(Met)、苯丙氨酸(Phe)、色氨酸(Try)、赖氨酸(Lys)和缬氨酸(Val)。20种氨基酸的分类,主要是根据R基来区分的。早些年根据R基的结构把氨基酸分为脂肪族、芳香族和杂环族三类,其中脂肪族又分为中性(一氨基一羧基)、酸性(一氨基二羧基)和碱性(二氨基一羧基)氨基酸。近年来都按R基的极性来区分氨酸的种类。
对于含有一个氨基和一个羧基的α–氨基酸来说,在中性溶液中或固体状态下,是以中性分子的形式还是以两性离子的式存在呢?许多实验证明主要是以两性离子的形式存在。
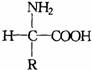
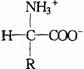
中性分子形式 两性离子形式
氨基酸由于含有氨基和羧基,因此在化学性质上表现为是的一种兼有弱碱和弱酸的两性化合物。氨基酸在溶液中的带电状态,会随着溶液的pH值而变化,如果氨基酸的净电荷等于零,在外加电场中不发生向正极或负极移动的现象,在这种状态下溶液的pH值称为其等电点,常用pI表示。由于各种氨基酸都有特定的等电点,因此当溶液的pH值低于某氨基酸的等电点时,则该氨基酸带净正电荷,在电场中向阴极移动。若溶液的pH值高于某氨基酸的等电点时,则该氨基酸带净负电荷,在电场中向阳极移动。氨基酸等电点的计算方法:对于单氨基单羧基的氨基酸,其等电点是pK1和Pk2的算术平均值,即从pI=1/2(pK1+pK2)公式中求得;对于含有3个可解离基团的氨基酸来说,只要依次写出它从酸性经过中性至碱性溶溶解高过程中的各种离子形式,然后取两性离子两侧的pK值的算术平均值,即可得其pI值。例如Asp解离时,有3个pK值,在不同pH条件下可以有4种离子形式,如下图所示。
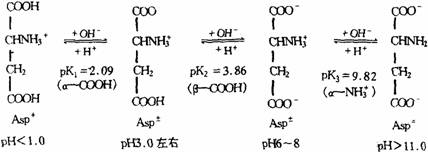
在等电点时,两性离子形式主要是Asp+,因此Asp的pI=1/2(pK1+pK2)=1/2(2.09+3.86)=2.98。同样方法可以求得其他含有3个pK值的氨基酸的等电点。各种氨基酸在可见光区都没有光吸收,而在紫外光区仅色氨酸、酪氨酸和苯丙氨酸有吸收能力。其中色氨酸的最大吸收波长为279nm,酪氨酸的最大吸收波长为278nm,苯丙氨酸的为259nm。利用紫外光法可以测定这些氨基酸的含量。蛋白质在280nm的紫外光吸收绝大部分是由色氨酸和酪氨酸所引起的。因此测定蛋白质含量时,用紫外分光光度法测定蛋白质在280nm的光吸收值是一种既简便而又快速的方法。
2.蛋白质的化学结构与空间结构
组成蛋白质的氨基酸,是借助肽键连接在一起的。肽键是由一个氨基酸分子中的α氨基与相邻的另一个氨基酸分子中α–羧基,通过失水缩合而成,这样连起来的氨基酸聚合物叫做肽。肽链上的各个氨基酸,由于在相互连接的过程中丢失了α–氨上的H和α–羧基上的OH,被称之为氨基酸残基。在多肽链一端氨基酸含有一个尚未反应的游离氨基(一NH2),称为肽链的氨末端氨基酸或N末端氨基酸;另一端的氨基酸含有一个尚未反应的游离羧基(-COOH),称为肽链的羧基末端氨基酸或C末端氨基酸。一般表示多肽链时,总是把N末端写在左边,C末端写右边。合成肽链时,合成方向是从N末端开始,逐渐向C末延伸。
各种蛋白质分子都有特定的空间结构,即构象。
蛋白质的一级结构:又称初级结构或化学结构,是指组成蛋质分子的多肽链中氨基酸的数目、种类和排列顺序,多肽链的数目,同时也包括链内或键间二硫键的数目和位置等。蛋白质分子的一级结构是由共价键形成的,肽键和二硫键都属于共价键。肽键是蛋白质分子中氨基酸连接的基本方式,形成共价主链。二硫键(-S-S)是由两个半胱氨酸(残基)脱氢连接而成的,是连接肽链内或肽链间的主要化学键。二硫键在蛋白质分子中起着稳定肽链空间结构的作用,往往与生物活力有关。二硫键被破坏后,蛋白质或多肽的生物活力就会丧失。蛋白质结构中,二硫键的数目多,蛋白质结构的稳定性就越强。在生物体内起保护作用的皮、角、毛发的蛋白质中,二硫键最多。
蛋白质的二级结构:是指多肽链本身折叠和盘绕方式,是指蛋白质分子中的肽链向单一方向卷曲而形成的有周期性重复的主体结构或构象。这种周期性的结构是以肽链内或各肽链间的氢键来维持。常见的二级结构有α–螺旋、β–折叠、β–转角等。例如动物的各种纤维蛋白,它们的分子围绕一个纵轴缠绕成螺旋状,称为α–螺旋。相邻的螺旋以氢键相连,以保持构象的稳定。指甲、毛发以及有蹄类的蹄、角、羊毛等的成分都是呈α–螺旋的纤维蛋白,又称α–角蛋白。β–折叠片是并列的比α–螺旋更为伸展的肽链,互相以氢铸连接起来而成为片层状,如蚕丝、蛛丝中的β–角蛋白。
蛋白质的三级结构:是指在二级结构的基础上,进一步卷曲折叠,构成一个很不规则的具有特定构象的蛋白质分子。维持三级结构的作用力主要是一些所谓弱的相互作用,即次级键或称非共价键,包括氢键、盐键、疏水键和范德华力等。盐键又称离子健,是蛋白质分子中正、负电荷的侧链基团互相接近,通过静电吸引而形成的,如羧基和氨基、胍基、咪唑基等基团之间的作用力。疏水键是多肽链上的某些氨基酸的疏水基团或疏水侧链(非极性侧链)由于避开水而造成相互接近、粘附聚集在一起。它在维持蛋白质三级结构方面占有突出地位。范德华引力是分子之间的吸引力。此外二硫键也对三级结构的构象起稳定作用。
具有三级结构的球蛋白是一类比纤维蛋白的构象更复杂的蛋白质。肽链也有α–螺旋、β–折叠片等构象单元,这些构象单元之间由肽链中不规则卷曲的肽段相连接,使整个肽铸折叠成近乎球状的不规则形状。酶、多种蛋白质激素、各种抗体以及细胞质和细胞膜中的蛋白质都是球蛋白。和纤维蛋白不同,球蛋白的表面富有亲水基团,因此都能溶于水。
蛋白质的四级结构:是由两条或两条以上的具有三级结构的多肽聚合而成特定构象的蛋白质分子。构成功能单位的各条肽链,称为亚基,一般地说,亚基单独存在时没有生物活力,只有聚合成四级结构才具有完整的生物活性。例如,磷酸化酶是由2个亚基构成的,谷氨酸脱氢酶是由6个相同的亚基构成的,血红蛋白是由4个不同的亚基(2个α肽链,2个β链)构成的,每个链都是一个具三级结构的球蛋白。亚基聚合成四级结构,是通过分子表面的一些次级键,主要是盐键和氢键结合而联系在一起的。有些蛋白质分子只有一、二、三级结构,并无四级结构,如肌红蛋白、细胞色素C、核糖核酸酶、溶菌酶等。另一些蛋白质,则一、二、三、四级结构同时存在,如血红蛋白、过氧化氢酶、谷氨酸脱氢酶等。
3.蛋白质的性质及生物学功能
蛋白质是由许多氨基酸分子组成的,分子量很大。所以它有的性质与氨基酸相同,有的性质又与氨基酸不同,如胶体性质、变构作用和变性作用等。
(1)胶体性质:蛋白质分子量很大,容易在水中形成胶体粒,具有胶体性质。在水溶液中,蛋白质形成亲水胶体,就是在胶体颗粒之外包含有一层水膜。水膜可以把各个颗粒相互隔开,所以颗粒不会凝聚成块而下沉。
(2)变构作用:含2个以上亚基的蛋白质分子,如果其中一个亚基与小分子物质结合,那就不但该亚基的空间结构要发生变化,其他亚基的构象也将发生变化,结果整个蛋白质分子的构象乃至活性均将发生变化,这一现象称为变构或别构作用。例如,某些酶分子可以和它所催化的最终产物结合,引起变构效应,使酶的活力降低,从而起到反馈抑制的效果。
(3)变性作用:蛋白质在重金属盐(汞盐、银盐、铜盐等)、酸、碱、乙醛、尿素等的存在下,或是加热至70-100℃,或在X射线、紫外线的作用下,其空间结构发生改变和破坏,从而失去生物学活性,这种现象称为变性。变性过程中不发生肽键断裂和二硫键的破坏,因而不发生一级结构的破坏,而主要发生氢键、疏水键的破坏,使肽链的有序的卷曲、折叠状态变为松散无序。
种类繁多的蛋白质具有多种多样的生物学功能,归纳起来主要具有下列5个方面:(1)作为酶,蛋白质具有催化功能。(2)作为结构成分,它规定和维持细胞的构造。(3)作为代谢的调节者(激素或阻遏物),它能协调和指导细胞内的化学过程。(4)作为运输工具,它能在细胞内或者透过细胞膜传递小分子或离子。(5)作为抗体,它起着保护有机体,防御外物入侵的作用。蛋白质是一切生命现象不可缺少的,即使像病毒、类病毒那样以核酸为主体的生物,也必须在它们寄生的活细胞的蛋白质的作用下,才能表现出生命现象。
在中学生物学教学大纲中,已简单介绍过蛋白质、核酸、ATP的结构和功能、酶和酶特性以及蛋白质的生物合成等知识,根据国际生物学奥林匹克竞赛纲要和全国中学生生物学竞赛大纲(试行)的要求,有关生物化学的内容在竞赛中经常要用到的一些知识,还必须在原有高中生物基础上加以充实和提高。
例1 甲、乙两个细胞分别在0.25M和0.35M的蔗糖溶液中发生初始质壁分离,若该两个细胞相邻,试问:
(1)在25℃条件下细胞间水分的移动方向?
(2)甲、乙两细胞的水势(ψw),渗透势(ψs)和压力势(ψp)分别是多少?
分析 解答此题,必须明确以下两个隐含条件,即(1)当细胞处于初始质壁分离时,细胞液浓度等于外液浓度;(2)细胞出现质壁分离以后,压力势(ψp)=0,细胞的水势(ψw)等于细胞的渗透势(ψs)。运用公式P=iCRT,即可求出细胞的渗透势(即水势),再根据甲、乙两细胞水势的大小来判断水分移动的方向。
解:依据公式P=iCRT,ψs=p,则
甲细胞的ψs=-iCRT=-1×0.25×0.082×(273+25)=-6.11(大气压)=-6.11×1.013=-6.19(巴)
乙细胞的ψs=-iCRT=-1×0.35×0.082×(273+25)=-8.56(大气压)=-8.56×1.013=-8.67(巴)
∵ψp=0 ∴ψs=ψw
故:甲细胞的ψw=-6.19巴,乙细胞的ψw=-8.67巴,乙细胞的水势低于甲细胞,所以水分从甲细胞向乙细胞移动。
例2 对下列说法中,正确的是
A 一个细胞放入某一浓度的溶液中时,若细胞液浓度与外界溶液的浓度相等,则体积不变。
B 若细胞的ψp=-ψs,将其放入某一溶液中时,则体积不变。
C 若细胞的ψw=ψs,将其放入纯水中,则体积变大。
D 有一充分饱和的细胞,将其放入比细胞液浓度低50倍的溶液中,则体积不变。
分析 A 除了处于初始质壁分离的状态的细胞之外,当细胞内液浓度与外液浓度相等时,由于细胞ψwp的存在,因而细胞水势高于外液水势而发生失水,体积会变小。
B 由于细胞ψw=0,则把该细胞放入任意溶液时,都会失水,体积变小。
C 当细胞的ψw=ψs时,将其放入纯水中,由于ψp=0,而ψs为一负值,故此时细胞吸水,体积变大。
D 充分胞和的细胞ψw=0,稀释溶液的ψw<0,所以该细胞会失水,体积变小。
(五)植物的成花生理
1.春化作用
低温对植物成花的促进作用称为春化作用。某些植物在其个体发育的某一个时期对低温有特殊要求,只有经低温的处理后,才能形成花芽。如冬小麦一般在秋季播种,冬前经历一段营养生长,经受低温后,于第二年夏初开花结实。冬小麦春播不抽穗,是因为未能满足它对低温的要求。如将萌动的种子放在0-5℃的低温中,经过30-50天,就可以春播而正常开花结实。一年生的冬性禾谷类作物、二年生的植物如甜菜、萝卜和大白菜以及某些多年生草本植物如牧草,都有春化现象。春化过程中感受低温的部位是芽内的分生组织,其作用只能随着细胞的分裂传递给子细胞。
2.植物的光周期诱导
(1)植物感受光周期的部位和时期
试验证明,植物感受光周期刺激的部位是叶片而不是生长点。如将短日植物菊花做如下处理:a.叶片在短日照下,生长点处在长日照;b.叶片进行长日照处理,生长点处在短日照下。结果a处理开了花,而b处理不开花。对光周期反应敏感的植物,如苍耳,只要有一片叶子、甚至半片叶子获得所需要的光周期即能使植物开花。萝卜、日本牵牛也如此。
植物对光周期的反应与叶龄有关,一般讲子叶无感光能力,幼嫩叶片的感受能力很小,充分展开的成熟叶片感受能力最强,老叶也失去感受能力。不同植物开始对光周期表现敏感的年龄也不同,如大豆、日本牵牛是在子叶伸展期,水稻在5-7叶时期,红麻则在6叶期。
(2)光周期中光与黑暗的意义
许多试验证明,对于短日照植物,在光周期中真正起诱导作用的是一定长度的黑暗,只要暗期达到临界夜长,不管先期的长短,短日植物均能开花。(如下图所示)
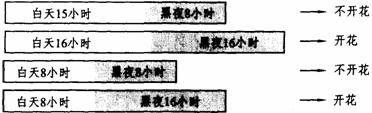
长日和短日或长夜或短夜影响苍耳开花的图解
对于长日植物来讲,只要暗期不超过临界夜长,无论光期的长短,也都可使之开花。可以这样讲,在光周期诱导中实际上是长夜诱导短日植物开花,抑制长日植物开花。对于植物的开花来讲,暗期比光照更重要。短日植物在超过一定的暗期长度时开花,长日植物则是在短于一定的暗期长度时开花。这也可由闪光实验得到证实(如下图所示)。

长日照的暗间断和长夜的光间断对长日植物和短日植物影响的图解
以上实验证明,用闪光中断长夜,尽管暗期的长度不变,却使长日植物开了花,而短日植物的开花受到了抑制;用黑暗中断长日,尽管光期变短,但却仍然是长日植物开花,短日植物不开花。充分说明了一定长度的连续暗期在花诱导中的重要性。
(3)光敏色素在成花中的作用
植物体中有一种光敏色素。这种色素在植物体内以两种状态存在,一种是吸收红光的状态(Pr),吸收峰为660nm;另一种是吸收远红光的状态(Pfr),吸收峰为730nm。Pr吸收红光后则转变成Pfr,而Pfr吸收远红光后则转变成Pr。Pfr是具有生理活性的形式。当Pfr与某些物质形成复合物后,则引起成花刺激物的合成。所以光敏色素在成花反应中的作用在于感受光,并诱导成花刺激物的形成。
光敏色素除了作为诱导植物开花的光受体外,还在许多光调节过程如需光种子萌发和光形态建成反应中起光受体作用。
湖北省互联网违法和不良信息举报平台 | 网上有害信息举报专区 | 电信诈骗举报专区 | 涉历史虚无主义有害信息举报专区 | 涉企侵权举报专区
违法和不良信息举报电话:027-86699610 举报邮箱:58377363@163.com